
rexresearch
DNA BioComputer Reprogramming
http://www.psychicchildren.co.uk/4-3-RussianDNAResearch.html
Russian DNA Research
Russian biophysicist and molecular biologist Pjotr Garjajev and his colleagues have been carrying out cutting-edge research the more esoteric nature of DNA. They simply did not believe that 90% of our DNA is ‘Junk DNA’. From the German book Vernetzte Intelligenz by Grazyna Fosar and Franz Bludorf (summarised and translated by Baerbel):
The latest research explains phenomena such as clairvoyance, intuition, spontaneous and remote acts of healing, self healing, affirmation techniques, unusual light-auras around people (namely spiritual masters), mind's influence on weather-patterns and much more. The Russian scientists also found out that our DNA can cause disturbing patterns in the vacuum, thus producing magnetized wormholes! Wormholes are the microscopic equivalents of the so-called Einstein-Rosen bridges in the vicinity of black holes (left by burned-out stars). These are tunnel connections between entirely different areas in the universe through which information can be transmitted outside of space and time. The DNA attracts these bits of information and passes them on to our consciousness...
Russian researcher Dr.Vladimir Poponin put DNA in a tube and beamed a laser through it. When the DNA was removed, the laser light continued spiralling on its own, like it would through a crystal! This effect is called ‘Phantom DNA Effect’.
It is surmised that energy from outside of space and time still flows through the activated wormholes after the DNA was removed. The side effect encountered most often in hyper communication and also in human beings are inexplicable electromagnetic fields in the vicinity of the persons concerned. Electronic devices like CD players and the like can be irritated and cease to function for hours. When the electromagnetic field slowly dissipates, the devices function normally again. Many healers and psychics know this effect from their work.
Chapter 9
The Mysterious DNA
Junk DNA
While western science invested in the International Human Genome Project focusing on the 5% of the encoding triplets of DNA, in the Soviet Union in 1990 a group of Russian scientists of the Russian Academy of Sciences was formed to study the complete human genome. This research was led by Dr. Pjotr Garjajev, member of the Russian Academy of Sciences as well as the Academy of Sciences in New York. The Russian research was taking a wide angle and held an open view in their studies. The research team included bio physicists, molecular biologists, embryologists and even linguistic experts. Their research revealed that the supposed junk DNA that has been completely neglected and forgotten by western mainstream science, was no redundant leftover of evolution at all. Linguistic studies revealed that the sequencing of the codons of the non-coding DNA follow the rules of some basic syntax. There is a definite structure and logic in the sequence of these triplets, like some biological language. Research further revealed that the codons actually form words and sentences just like our ordinary human language follows grammar rules.
Scientists have conducted much research on the origins of human languages and the origins of the grammatical rules that are so essential to all human languages; however they have always failed to find the source. But now for the first time in history the origins of language may be surprisingly attributed to DNA. The language of the genes is much, much older than any human language that was ever uttered on this globe. It is even conceivable that the DNA grammar itself served as the blueprint for the development of human speech.
Whereas the western Human Genome Project deciphered the ‘machine language’ code of the DNA molecule, the structure of the DNA ‘bits’ formed by the sequence of nucleotides, Russian scholars discovered the higher level language present in DNA. Another amazing fact that Garjajev’s group discovered was that the DNA is by no means a closed book of life. He discovered that the text of the DNA book can be altered. The codons of the DNA string can be rearranged in different sequences. In other words the software of the human genome our DNA molecule can be reprogrammed! Research revealed that the triplets in the DNA string are able to exchange places.
Since the DNA was found to have a syntax and semantics akin to our human languages, it indicated that our currently restricted understanding of DNA serving only for the coding of the reproduction of proteins for the chemical make up of an organism, is only half of the story.
When in vitro DNA in test tubes was exposed to coherent laser light, the laser light spiralled along the DNA helix as if it was guided by the structure of the DNA molecule. The most amazing effect was noticed when the DNA itself was removed and the laser light kept spiralling! The vacuum of the space that was just previously occupied by the DNA had changed and something caused the laser light to keep spiralling. These effects have been measured and remained for quite some time after the DNA was removed. The effect is now becoming well known as the DNA phantom effect. Vladimir Poponin and his team of Russian Academy of Sciences repeated the work of Garjajev at the Hearthmath Institute in the U.S.A. Poponin concluded again that a field structure was formed in the physical vacuum even when the original DNA was removed. We’ve seen similar examples of vacuum changes before that could be attributed to torsion fields. (1)
DNA programming
The most astonishing experiment that was performed by Garjajev’s group is the reprogramming of the DNA codon sequences using modulated laser light. From their discovered grammatical syntax of the DNA language they were able to modulate coherent laser light and even radio waves and add semantics (meaning) to the carrier wave. In this way they were able to reprogram in vivo DNA in living organisms, by using the correct resonant frequencies of DNA. The most impressive discovery made so far is that spoken language can be modulated to the carrier wave with the same reprogramming effect. Now this is a baffling and stunning scientific discovery! Our own DNA can simply be reprogrammed by human speech, supposing that the words are modulated on the correct carrier frequencies!
Whereas western science uses complicated bio chemical processes to cut and paste DNA triplets in the DNA molecule, Russian scientist use modulated laser light to do exactly the same thing. The Russians have proven to be very successful in repairing damaged DNA material in vivo!
Laser light therapies based on Garjajev’s findings are already applied in some European academic hospitals with success on various sorts of skin cancer. The cancer is cured without any remaining scars.
(2)...
When Our Cell Phones Chat with Our Genes
By Grazyna Fosar and Franz Bludorf
"...In truth, DNA is not just a blueprint for constructing the body; it is also a storage medium for optical information as well as an organ for communication.
The German biophysicist Fritz-Albert Popp spent many years studying biophoton emission, i.e., the light that the body naturally emits. Popp’s studies indicate that DNA is one of the main suppliers of biophotons. Through comprehensive studies, he discovered that DNA is a harmonic oscillator – an oscillating system with its own particular frequency, or resonating frequency. Using the length of the DNA molecule (about two meters, when stretched out), and the known speed of light, DNA’s own frequency is calculated as about 150 megahertz...
The principles have been worked out by Dr. Piotr P. Garjajev and his colleagues at the Russian Academy of Science in Moscow.
Genes Speak Our Language
Garjajev’s findings go far beyond Popp’s: According to Garjajev’s studies, DNA is not only the transmitter and receiver of electromagnetic radiation (in the form of energy), but it also absorbs information contained in the radiation and interprets it further. Thus, DNA is an extremely complex interactive optical biochip. In our book Vernetzte Intelligenz (Cross-Linked Intelligence) we wrote the following on this subject:
“For example, today we speak almost matter-of-factly about the “genetic code,” that is, about a systematic codification of information. However, genetics to date has gotten stuck at this point, and has relied exclusively on chemistry to finish the work, without even once calling in language experts.
“In Moscow, things are different. There, the genetic code was…additionally subjected to thorough examination by linguists. (Linguistics is the science of the structure and formation of languages). When studying a language, people investigate regularities such as syntax (rules for building words from letters), semantics (the study of the content meaning of words), and rules of grammar.
“When this scientific knowledge is applied to the genetic code, it can be seen that this code follows the same rules as our human languages.
“It should be noted: not the rules of a particular language (in this case, for example, the Russian language), but rather rules at a fundamental level where common features reside among all existing languages of mankind. Thus, the structure of the genetic code can be connected with every existing human language.
“Scientists have been seeking the ancestral human language for centuries – Piotr Garjajev and his colleagues may have found it.”
http://www.bethcoleman.net/intelligenz.html
Quantum Physics in for Radical Revision - DNA
From the book, "Vernetzte Intelligenz"
by
Grazyna Fosar und Franz Bludorf
The human DNA is a biological Internet and superior in many aspects to the artificial one. The latest Russian scientific research directly or indirectly explains phenomena such as clairvoyance, intuition, spontaneous and remote acts of healing, self healing, affirmation techniques, unusual light/auras around people (namely spiritual masters), mind´s influence on weather patterns and much more. In addition, there is evidence for a whole new type of medicine in which DNA can be influenced and reprogrammed by words and frequencies WITHOUT cutting out and replacing single genes.
Only 10% of our DNA is being used for building proteins. It is this subset of DNA that is of interest to western researchers and is being examined and categorized. The other 90% are considered "junk DNA." The Russian researchers, however, convinced that nature was not dumb, joined linguists and geneticists in a venture to explore those 90% of "junk DNA." Their results, findings and conclusions are simply revolutionary!
According to them, our DNA is not only responsible for the construction of our body but also serves as data storage and in communication. The Russian linguists found that the genetic code, especially in the apparently useless 90%, follows the same rules as all our human languages. To this end they compared the rules of syntax (the way in which words are put together to form phrases and sentences), semantics (the study of meaning in language forms) and the basic rules of grammar.
They found that the alkalines of our DNA follow a regular grammar and do have set rules just like our languages. So human languages did not appear coincidentally but are a reflection of our inherent DNA.
The Russian biophysicist and molecular biologist Pjotr Garjajev and his colleagues also explored the vibrational behavior of the DNA. [For the sake of brevity I will give only a summary here. For further exploration please refer to the appendix at the end of this article.] The bottom line was: "Living chromosomes function just like solitonic/holographic computers using the endogenous DNA laser radiation." This means that they managed for example to modulate certain frequency patterns onto a laser ray and with it influenced the DNA frequency and thus the genetic information itself.
Since the basic structure of DNA-alkaline pairs and of language (as explained earlier) are of the same structure, no DNA decoding is necessary. One can simply use words and sentences of the human language! This, too, was experimentally proven! Living DNA substance (in living tissue, not in vitro) will always react to language-modulated laser rays and even to radio waves, if the proper frequencies are being used. This finally and scientifically explains why affirmations, autogenous training, hypnosis and the like can have such strong effects on humans and their bodies. It is entirely normal and natural for our DNA to react to language. While western researchers cut single genes from the DNA strands and insert them elsewhere, the Russians enthusiastically worked on devices that can influence the cellular metabolism through suitable modulated radio and light frequencies and thus repair genetic defects.
Garjajev´s research group succeeded in proving that with this method chromosomes damaged by x-rays for example can be repaired. They even captured information patterns of a particular DNA and transmitted it onto another, thus reprogramming cells to another genome. So they successfully transformed, for example, frog embryos to salamander embryos simply by transmitting the DNA information patterns! This way the entire information was transmitted without any of the side effects or disharmonies encountered when cutting out and re-introducing single genes from the DNA.
This represents an unbelievable, world-transforming revolution and sensation! All this by simply applying vibration and language instead of the archaic cutting-out procedure! This experiment points to the immense power of wave genetics, which obviously has a greater influence on the formation of organisms than the biochemical processes of alkaline sequences.
Esoteric and spiritual teachers have known for ages that our body is programmable by language, words and thought. This has now been scientifically proven and explained. Of course the frequency has to be correct. And this is why not everybody is equally successful or can do it with always the same strength. The individual person must work on the inner processes and maturity in order to establish a conscious communication with the DNA. The Russian researchers work on a method that is not dependent on these factors but will ALWAYS work, provided one uses the correct frequency.
But the higher developed an individual´s consciousness is, the less need is there for any type of device! One can achieve these results by oneself, and science will finally stop to laugh at such ideas and will confirm and explain the results. And it doesn´t end there.
The Russian scientists also found out that our DNA can cause disturbing patterns in the vacuum, thus producing magnetized wormholes! Wormholes are the microscopic equivalents of the so-called Einstein-Rosen bridges in the vicinity of black holes (left by burned-out stars). These are tunnel connections between entirely different areas in the universe through which information can be transmitted outside of space and time. The DNA attracts these bits of information and passes them on to our consciousness. This process of hypercommunication is most effective in a state of relaxation.
Stress, worries or a hyperactive intellect prevent successful hypercommunication or the information will be totally distorted and useless. In nature, hypercommunication has been successfully applied for millions of years. The organized flow of life in insect states proves this dramatically. Modern man knows it only on a much more subtle level as "intuition." But we, too, can regain full use of it...
All informations are from the book "Vernetzte Intelligenz" von Grazyna Fosar und Franz Bludorf, ISBN 3930243237, summarized and commented by Baerbel. The book is unfortunately only available in German so far. You can reach the authors here: [www.fosar-bludorf.com] [2]; Transmitted by Vitae Bergman [www.ryze.com/view.php?who=vitaeb]
http://www.fosar-bludorf.com/archiv/biochip_eng.htm
Revolutionary Results of Modern Genetics
by Grazyna Fosar and Franz Bludorf
The Biological Chip in our Cells
Independent of the biochemical function as a protein producer the DNA is a complicated electronic biological chip that communicates with its environment, as latest research from Russia found out.
In the year 1990 a group of scientists got together in Moscow, for whom the study of the human Genoms was too much reduced exclusively to biochemistry. They had recognized that by this viewpoint, which is based rather on orthodox dogmatism than on objective scientific realizations a lot of information remains hidden to us.
Highly-qualified scientists belong to this group, to a large extent from the Russian Academy of Sciences. Beside physicists of the renowned Lebedev institute also molecular biologists participate, bio physicists, geneticists, embryologists and linguists. Director of the project is Dr. Pjotr Garjajev, a bio physicist and molecular biologist. He is member of the Russian Academy of Sciences as well as of the Academy of Sciences in New York.
In the eight years since establishment of the project the Muscovite group came to revolutionary realizations, which let our understanding of the DNA and the human genetics appear in a completely new light.
For example we speak today nearly naturally of the »genetic code«, thus of a systematic information coding. But the past genetics stopped here and settled the remainder of the work exclusive with the help of chemistry, instead of consulting also language experts.
Differently in Moscow. Here, as already mentioned, the genetic code was submitted an exact investigation by linguists too.
Linguistics is the science of the structure of languages. It investigates thereby not only the natural languages, which developed in the individual countries and cultures, but also artificial languages, which are used for example for programming computers and which were developed in the past decades systematically using linguistic realizations.
One examines semantics (theory of the meaning of the words) and language regularities like the syntax (rules for the setting up of words from letters), as well as the bases of the grammar.
If one uses these scientific realizations on the genetic code, then one recognizes that this code follows the same rules as our human languages.
Mind you: not the rules of a certain language (in this case e.g. the Russian one), but on such a fundamental level, where all existing languages of mankind have comparable structures. So it is possible to set the structure of the genetic code in relationship with each existing language of mankind.
For centuries scientists looked for the human original language - Pjotr Garjajev and his coworkers possibly found it.
We must turn around the relations: the structure of the DNA does not correspond to the human language structure, but the human languages follow the genetic code in their structure the rules! DNA and genetic code existed already for a long time, before first humans spoke an articulated word for the first time! Every human languages developed since that time followed the basic pattern, already existing in the structure of the genetic code.
You should not misunderstand this realization: It does not concern here an orthodox materialistic conception of the world, according to which the ability for speaking would be only a secondary effect of proteins, which are put on in some genes. The arrangement of the elementary bases in the DNA follows a grammar, an immaterial plan, which is similar to the structure of our languages.
The fact that no physical procedure is concerned here is proved by the next discovery of Garjajev's team: The analogy between the structure of the DNA and the human language is most pronounced just in the parts of the giant molecule, which are not used for protein synthesis!
For a long time one knows that only about 10 per cent of the DNA molecule are used for setting up genes. The remaining 90 per cent have a function unknown to classical science and were designated so far as »silent DNA«.
Garjajev's realization thus is a revolution for the entire area of genetics. Examining only the well-known genes calling the remainder »silent DNA«, you will miss the most important facts! It is paradoxical: just the »silent DNA« - figurativy spoken - speaks a language!
In various experiments the Muscovites group could prove that these extensive codes in the DNA are not used by any means for the synthesis of a so far unknown quantity of components of our body, as it is the case with the genes. This code is rather actually used for communication, more exactly - for hypercommunication.
Hypercommunication is a data exchange on DNA level using genetic code. Since this code possesses a structure, which is the basis of all human languages, also higher information may be transported, which is able to come up to human consciousness and to be interpreted there.
Garjajev and its colleague continued still another step. They analyzed the vibration response of the DNA and found out that it follows quite complicated laws, which are however well known in the physics for a long time.
Those are the laws of nonlinear waveform-shaping, known since center of 19 century as so-called Soliton waves. They are known from observations, but so complicated that they may be calculated only with modern computers. Soliton waves are temporally extraordinarily stable and may store information in this way for a long time.
Summarizing all these realizations, one comes to a perfectly new form of the genetic engineering, possibly even to a new gene therapy.
Concerning this Pjotr Garjajev writes: »The majority tries to understand the principles of the DNA biological computer by appointing oneself exclusively to the DNA Watson Crick Chargaff rules: A-T, G-C. That is correct, but it is so not enough! The DNA chromosomale continuum in living systems has wave attributes, which lets us derive the unknown, a computer-similar program for the setting up of the organisms. The well-known genetic code is a code for protein synthesis and nothing further. Chromosomes in vivo work as solitonic holographic computers under use of the endogenous DNA laser radiation.«
This sounds like science of the 21st century and probably is. But don't forget: Garjajevs statements are founded scientifically in theory and experiment.
The consequences of these realizations are as incomprehensible as simple and logical: If one modulates a laser beam by a frequency sample, then one may affect with this the information of the DNA waves and so the genetic information itself.
For this one does not even need to decode the language of the pairs of bases in laborious work, in order to formulate from it artificially genetic information, but one can use quite easily words and sentences of the human language! The bases of the language structures are, as we in now know, the same.
Also this astonishing conclusion the Muscovites group of researchers could prove already experimentally. DNA substance in vivo (i.e. in the living fabric, not in the test tube) reacts to language-modulated laser light, even to radio waves, if one keeps the correct resonant frequencies.
In this way unknown possibilities are opened to the medicine. One may design devices, with which through suitably modulated radio or light radiation cell metabolism may be affected, even the repair of genetic defects is possible, without all the risks and side effects of the classical-biochemical proceeding.
Garjajevs group of researchers could already prove that with this method chromosomes may be repaired, which were damaged e.g. by x-rays. The effects on medical therapy possibilities of the coming century are immense: one can develop devices for new, subtle cancer therapy, also for the treatment of aids and for the slowing down of the aging process.
Already today devices are in use also in German university clinics, with whose assistance cancer patients are exposed to frequency-modulated magnetic field irradiation. The results are promising.
We see here that the objectives of the Muscovites researchers deviate from those of the western human Genome Project in principle. While in the western science the trend is to develop new chemical medicines from as much as possible items of information from the genes - a procedure, which is however not free from substantial risks, a potential giant business -, the Russian scientists have a rather holistic understanding of the DNA leading to the development of therapy devices, which may replace some expensive and dangerous medicine in the long term.
Quite beside from the new wave theory of the genetic code still some further interesting facts follow. For example one knows for a long time that almost any bodily function, particularly also in the metabolism and in the hormone production, can be affected by suggestive strength of the spoken word, although they run perfectly autonomously, thus under elimination of the conscious will, whereupon the impact of the medical hypnosis is based. These facts are well-known, however could not be explained so far scientifically.
The medical model of the psycho neuro immunology led back the effect of hypnotic suggestions so far exclusively to control mechanisms in the brain, particularly in the regions, which are assumed to contain subconscious layers.
Now it looks that it is much simpler: the DNA is able to react directly to the spoken word.
Also different therapy procedures, whose impact was inexplicable so far, as for instance the Chinese acupunkture, may be explained with help of the DNA wave theory in Garjajev's opinion scientifically.
If we summarize the research results of Professor Popp and Professor Garjajev, then a remarkable connection results: Light actually represents an important factor in the power supply of our hereditary molecule, the DNA. It provides healthy functioning of all procedures in our cells. However it cannot form a complete replacement for material food alone.
The information, which will transfer via the light, is much more important. The DNA communicates in this way - perhaps with other organisms or with a superordinate plan - which a morphogenetic field, which could be proven by the research in Russia for the first time scientifically.
In this way the genetic information of each cell can employ comparisons of their actual condition with a specified condition each time and arrange possibly necessary repairs. This can prevent or at least stop diseases such as cancer or aids, in addition, slow down the age process.
The modern wave genetics is one of the key technologies for the coming millennium, and we can be strained, what science will discover in this area in the very near future.
I Speak of Dreams: Annals of Quackery: Hypnosis and Dyslexia
4. The press release also claims that hypnosis is a valid treatment for dyslexia based on a "scientific release" from a Russian scientist, biophysicist and molecular biologist Pjotr Garjajev, with the following claims"
"scientifically proven that affirmations along with meditation/ hypnosis (another term for meditation) will raise consciousness, well-being, and even change DNA
Garjajev has no citations in Pubmed. The claim for DNA modulation seems to come from the article entitled, "The Biological Chip in our Cells", by Grazyna Fosar and Franz Bludorf, published on their website, "German Magazine KonteXt reports on current developments within the ranges of border science and spirituality." There are a number of claims made, but no data to back up the claims. Remember, "Extraordinary claims demand extraordinary proof". We can reject the Fosar-Bludorf claims for lacking evidence...
http://www.homepages.ihug.co.nz/~sai/quant_biology.htm
Quantum Phenomena in Biology
1. Genetic code only half the story - DNA is a quantum mechanical biowave computer
Recent research has demonstrated that electromagentic signals are of key importance in the regulatory functioning of DNA . Part of it is based on ultraviolet luminence creating biophotons that have been experimentally demonstrated to be able to enhance metabolic reactions thousandfold (1). Another aspect is an electromagnetically mediated "language" for communication between DNA and the cells. Actually laser light generated in DNA, experimentally demonstrated by P.P. Garajev (2),(3) is a key element in this information transmission system.
Excerpt from:
Gariaev P., et al, "The DNA-wave Biocomputer" (3)
"...These assumptions produce a chromosome apparatus and fast wave genetic information channels connecting the chromosomes of the separate cells of an organism into a holistic continuum, working as the biocomputer, where one of the field types produced by the chromosomes, are their radiations. This postulated capability of such "laser radiations" from chromosomes and DNA, as will be shown, has already been demonstrated experimentally in Moscow, by the Gariaev Group. Thus it seems the accepted notions about the genetic code must change fundamentally, and in doing so it will be not only be possible to create and understand DNA as a wave biocomputer, but to gain from nature a more fundamental understanding of what information [Marcer in press] really is! For the Gariaev Group's experiments in Moscow and Toronto say that the current understanding of genomic information i.e. the genetic code, is only half the story [Marcer this volume]. "
References
1. Kaznacejev V. P., Michailova L. P. Ultraschwache Luminiszenz in interzellularen Interaktionen. Novosibirsk, Nauka, 1981 (in Russian.)
2. Garjajev P. P. Der wellengenetische Code. 1997, ISBN 5-7816-0022-1 (in Russian.)
3. Peter P. Gariaev, Boris I. Birshtein, Alexander M. Iarochenko, Peter J. Marcer, George G. Tertishny, Katherine A. Leonova, Uwe Kaempf, "The DNA-wave Biocomputer" at http://www.rialian.com/rnboyd/dna-wave.doc
The DNA-Wave Biocomputer
Peter P. Gariaev*, Boris I. Birshtein*, Alexander M. Iarochenko*, Peter J. Marcer**,
George G. Tertishny*, Katherine A. Leonova*, Uwe Kaempf ***.
* Institute Control of Sciences Russian Academy of Sciences,
Moscow, Russia gariaev@aha.ru, http://www.aha.ru/~gariaev, and
Wave Genetics Inc. 87 Scollard Street, Toronto, Ontario, Canada, M5R 1G4, gariaev@wavegenetics.com,
**53 Old Vicarage Green, Keynsham, Bristol, BS31 2DH, UK,
petermarcer@aikido.freeserve.co.uk,
http://www.bcs.org.uk/cybergroup.htm
*** Institut f. Klinische, Diagnostische und Differentielle Psychologie- Am Falkenbrunnen - D-01062 Dresden TU, Dresden, Germany, uwe@psy1.psych.tu-dresden.de
Abstract
This paper reports experimental work carried out in Moscow at the Institute of Control Sciences, Wave Genetics Inc. and theoretical work from several sources. This work changes the notion about the genetic code essentially. It asserts: -
1) That the evolution of biosystems has created genetic "texts", similar to natural context dependent texts in human languages, shaping the text of these speech-like patterns.
2) That the chromosome apparatus acts simultaneously both as a source and receiver of these genetic texts, respectively decoding and encoding them, and
3) That the chromosome continuum of multicellular organisms is analogous to a static-dynamical multiplex time-space holographic grating, which comprises the space-time of an organism in a convoluted form.
That is to say, the DNA action, theory predicts and which experiment confirms,
i) is that of a "gene-sign" laser and its solitonic electro-acoustic fields, such that the gene-biocomputer "reads and understands" these texts in a manner similar to human thinking, but at its own genomic level of "reasoning". It asserts that natural human texts (irrespectively of the language used), and genetic "texts" have similar mathematical-linguistic and entropic-statistic characteristics, where these concern the fractality of the distribution of the character frequency density in the natural and genetic texts, and where in case of genetic "texts", the characters are identified with the nucleotides, and ii) that DNA molecules, conceived as a gene-sign continuum of any biosystem, are able to form holographic pre-images of biostructures and of the organism as a whole as a registry of dynamical "wave copies" or "matrixes”, succeeding each other. This continuum is the measuring, calibrating field for constructing its biosystem.
Keywords: DNA, wave-biocomputer, genetic code, human language, quantum holography.
1. What Theory Predicts.
1.1 Introduction.
How did this new theory take shape? The principle problem of the creation of the genetic code, as seen in all the approaches [Gariaev 1994; Fatmi et al. 1990; Perez 1991: Clement et al. 1993; Marcer, Schempp 1996; Patel, 2000] was to explain the mechanism by means of which a third nucleotide in an encoding triplet, is selected. To understand, what kind of mechanism resolves this typically linguistic problem of removing homonym indefiniteness, it is necessary firstly to postulate a mechanism for the context-wave orientations of ribosomes in order to resolve the problem of a precise selection of amino acid during protein synthesis [Maslow, Gariaev 1994]. This requires that some general informational intermediator function with a very small capacity, within the process of convolution versus development of sign regulative patterns of the genome-biocomputer endogenous physical fields. It lead to the conceptualization of the genome's associative-holographic memory and its quantum nonlocality. These assumptions produce a chromosome apparatus and fast wave genetic information channels connecting the chromosomes of the separate cells of an organism into a holistic continuum, working as the biocomputer, where one of the field types produced by the chromosomes, are their radiations. This postulated capability of such "laser radiations" from chromosomes and DNA, as will be shown, has already been demonstrated experimentally in Moscow, by the Gariaev Group. Thus it seems the accepted notions about the genetic code must change fundamentally, and in doing so it will be not only be possible to create and understand DNA as a wave biocomputer, but to gain from nature a more fundamental understanding of what information [Marcer in press] really is! For the Gariaev Group's experiments in Moscow and Toronto say that the current understanding of genomic information i.e. the genetic code, is only half the story [Marcer this volume].
1.2 What experiment confirms, part one.
These wave approaches all require that the fundamental property of the chromosome apparatus is the nonlocality of the genetic information. In particular, quantum nonlocality/teleportation within the framework of concepts introduced by Einstein, Podolsky and Rosen (EPR) [Sudbery 1997; Bouwmeester et al.1997]. This quantum nonlocality has now, by the experimental work of the Gariaev Group, been directly related (i) to laser radiations from chromosomes, (ii) to the ability of the chromosome to gyrate the polarization plane of its own radiated and occluded photons and (iii) to the suspected ability of chromosomes, to transform their own genetic-sign laser radiations into broadband genetic-sign radio waves. In the latter case, the polarizations of chromosome laser photons are connected nonlocally and coherently to polarizations of radio waves. Partially, this was proved during experiments in vitro, when the DNA preparations interplaying with a laser beam (=632.8nm ), organized in a certain way, polarize and convert the beam simultaneously into a radio-frequency range. In these experiments, another extremely relevant phenomenon was detected: photons, modulated within their polarization by molecules of the DNA preparation. These are found to be localized (or "recorded") in the form of a system of laser mirrors' heterogeneities. Further, this signal can "be read out" without any essential loss of the information (as theory predicts [ Gariaev 1994; Marcer, Schempp 1996]), in the form of isomorphously (in relation to photons) polarized radio waves. Both the theoretical and experimental research on the convoluted condition of localized photons therefore testifies in favour of these propositions.
These independently research approaches also lead to the postulate, that the liquid crystal phases of the chromosome apparatus (the laser mirror analogues) can be considered as a fractal environment to store the localized photons, so as to create a coherent continuum of quantum-nonlocally distributed polarized radio wave genomic information. To a certain extent, this corresponds with the idea of the genome's quantum-nonlocality, postulated earlier, or to be precise, with a variation of it.
This variation says that the genetic wave information from DNA, recorded within the polarizations of connected photons, being quantum-nonlocal, constitutes a broadband radio wave spectrum correlated - by means of polarizations - with the photons. Here, the main information channel, at least in regard to DNA, is the parameter of polarization, which is nonlocal and is the same for both photons and the radio waves. A characteristic feature is, that the Fourier-image of the radio spectra is dynamic, depending essentially on the type of matter interrogated. It can therefore be asserted, that this phenomenon concerns a new type of a computer (and biocomputer) memory, and also a new type of EPR spectroscopy,namely one featuring photon-laser-radiowave polarization spectroscopy.The fundamental notion is, that the photon-laser-radiowave features of different objects (i.e. the Fourier-spectra of the radiowaves of crystals, water, metals, DNA, etc) are stored for definite but varying times by means of laser mirrors, such that the "mirror spectra" concern chaotic attractors with a complex dynamic fractal dynamics, recurring in time. The Gariaev Group experiments are therefore not only unique in themselves, they are a first example, that a novel static storage/recording environment (laser mirrors) exists, capable of directly recording the space-time atomic/molecular rotary dynamical behaviour of objects. Further the phenomena, detected by these experiments described in part two, establish the existence of an essentially new type of radio signal, where the information is encoded by polarizations of electromagnetic vectors. This will be the basis of a new type of video recording, and will create a new form of cinema as well.
Further experimental research has revealed the high biological (genetic) activity of such radio waves, when generated under the right conditions by DNA. For example, by means of such artificially produced DNA radiations, the super fast growth of potatoes (up to 1 cm per day) has been achieved, together with dramatic changes of morphogenesis resulting in the formation of small tubers not on rootstocks but on stalks. The same radiations also turned out to be able to cause a statistically authentic "resuscitation" of dead seeds of the plant Arabidopsis thaliana, which were taken from the Chernobyl area in 1987. By contrast, the monitoring of irradiations by polarized radio waves, which do not carry information from the DNA, is observed to be biologically inactive. In this sequence of experiments, additional evidence was also obtained in favour of the possibility of the existence of the genetic information in form of the polarization of a radio wave physical field. This supports the supposition that the main information channel in these experiments is the biosign modulations of polarizations mediated by some version of quantum nonlocality. A well known fact can therefore be seen in new light, namely, that the information biomacromolecules - DNA, RNA and proteins - have an outspoken capacity to optical rotatory dispersion of visible light and of circular dichroism. Similarly, the low molecular components of biosystems, such as saccharides, nucleotides, amino acids, porphyrins and other biosubstances have the same capacity; a capacity, which until now made little biological sense. Now, however, it supports, the contention that this newly detected phenomenon of quantized optical activity can be considered as the means by which the organism obtains unlimited information on its own metabolism. That is, such information is read by endogenous laser radiations of chromosomes, which, in their turn, produce the regulative ("semantic") radio emission of the genome biocomputer. Furthermore, the apparent inconsistency between the wavelengths of such radiations and the sizes of organisms, cells and subcell structures is abrogated, since the semantic resonances in the biosystems' space are realized not at the wavelength level, but at the level of frequencies and angles of twist of the polarization modes. This mechanism is the basis for the artificial laser-radio-wave vitro-in vivo scanning of the organism and its components.
However, chromosome quantum nonlocality as a phenomenon of the genetic information is seen as particularly important in multicellular organisms and as applying on various levels.
The 1-st level is that the organism as a whole. Here nonlocality is reflected in the capacity for regeneration, such that any part of the body recreates the whole organism, as, for example, in case of the worm Planaria. That is to say, any local limiting of the genetic information to any part of a biosystem is totally absent. The same concerns the vegetative reproduction of plants.
The 2nd level is the cellular level. Here it is possible to grow a whole organism out of a single cell. However with highly evolved animal biosystems, this will be a complex matter.
The 3rd level is the cellular-nuclear level. The enucleation of nuclei from somatic and sexual cells and the subsequent introduction into them of other nuclei does not impede the development of a normal organism. Cloning of this kind has already been carried out on higher biosystems, for example, sheep.
The 4th level is the molecular level: here, the ribosome "would read" mRNA not only on the separate codons, but also on the whole and in consideration of context.
The 5th level is the chromosome-holographic: at this level, a gene has a holographic memory, which is typically distributed, associative, and nonlocal, where the holograms "are read" by electromagnetic or acoustic fields. These carry the gene-wave information out beyond the limits of the chromosome structure. Thus, at this and subsequent levels, the nonlocality takes on its dualistic material-wave nature, as may also be true for the holographic memory of the cerebral cortex [ Pribram 1991; Schempp 1992; 1993; Marcer, Schempp 1997; 1998]
The 6th level concerns the genome's quantum nonlocality. Up to the 6th level, the nonlocality of bio-information is realized within the space of an organism. The 6th level has, however, a special nature; not only because it is realized at a quantum level, but also because it works both throughout the space of a biosystem and in a biosystems own time frame. The billions of an organism's cells therefore "know" about each other instantaneously, allowing the cell set is to regulate and coordinate its metabolism and its own functions. Thus, nonlocality can be postulated to be the key factor explaining the astonishing evolutionary achievement of multicellular biosystems. This factor says that bioinformatic events, can be instantaneously coordinated, taking place "here and there simultaneously", and that in such situations the concept of "cause and effect" loses any sense. This is of a great importance! The intercellular diffusion of signal substances and of the nervous processes is far too inertial for this purpose. Even if it is conceded that intercellular transmissions take place electro-magnetically at light speeds, this would still be insufficient to explain how highly evolved, highly complex biosystems work in real time [Gariaev 1994; Ho 1993]. The apparatus of quantum nonlocality and holography is in authors' view, indispensable to a proper explanation of such real time working. The 6th level therefore says, the genes can act as quantum objects, and that, it is the phenomenon of quantum non-locality/teleportation, that ensures the organism's super coherency, information super redundancy, super knowledge, cohesion and, as a totality or whole, the organism's integrity (viability).
Indeed it can be said that
this new understanding of biocomputers, constitutes a further
step in a development of computer technology in general. An
understanding that will bring about a total change of the
constituent basis of that technology, in the history of analogue
> to > digital > to > now, the figurative semantic
(nonlocal) wave computer or biocomputer. This biocomputer will
be based on new understanding of the higher forms of the DNA
memory, and the chromosome apparatus, as the recording,
storaging, transducing and transmitting system for genetic
information, that must be considered simultaneously both at the
level of matter and at the level of physical fields. The latter
fields, having been just studied, as showed experimentally in
this research, are carriers of genetic and general regulative
information, operating on a continuum of genetic molecules (DNA,
RNA, proteins, etc). Here, previously unknown types of memory
(soliton, holographic, polarization) and also the DNA molecule,
work both as biolasers and as a recording environment for these
laser signals. The genetic code, considered from such a point of
view, will be essentially different from today's generally
accepted but incomplete model. This, the wave-biocomputer model
asserts, only begins to explain the apparatus of protein
biosynthesis of living organisms, providing an important
interpretation for the initial stages within this new proposed
composite hierarchic chain of material and field, sign,
holographic, semiotic-semantic and, in the general case, of
figurative encoding and deciphering chromosome functions. Here
the DNA molecules, conceived as a gene-sign continuum of any
biosystem, are able to form pre-images of biostructures and of
the organism as a whole as a registry of dynamical "wave copies"
or "matrixes", succeeding each other. This continuum is the
measuring, calibrating field for constructing any biosystem.
1.3 Features of the Wave Model
Adleman
[1994], for example, has used the mechanism for fast and precise
mutual recognition between the DNA anti-parallels half-chains to
solve the "the travelling salesman's problem". However in the
wave model of biosystems, this is only one aspect of the
self-organization taking place. For here, as the experimental
evidence now confirms, the mutual recognition of one DNA anti
parallel half chain (+) by the other (-) concerns special super
persistent/resonant acoustic-electromagnetic waves or solitons.
Such DNA solitons have two connected types of memory. The first
is typical of the phenomenon discovered by Fermi-Pasta-Ulam
(FPU) [Fermi, 1972]. It concerns the capability of non-linear
systems to remember initial modes of energisation and to
periodically repeat them [Dubois 1992]. The DNA liquid crystals
within the chromosome structure form such a non-linear system.
The second is that of the DNA-continuum in an organism. Such
memory is an aspect of the genome's nonlocality. It is
quasi-holographic/fractal, and relates, as is the case for any
hologram or fractal, to the fundamental property of biosystems
i.e. to their ability to restore the whole out of a part. This
property is well known (grafting of plants, regeneration of a
lizard's tail, regeneration of a whole organism from the
oocyte). And a higher form of such a biological memory would be
a holographic (associative) memory of the brain cortex, i.e. of
its neural network [Pribram 1991; Schempp 1992; Marcer Schempp
1997, 1998; Sutherland 1999]. Such wave sign encoding/decoding
therefore, like DNA's ability to resolve "the traveling
salesman's problem", is, it can be hypothesized, an integral
part of DNA's computational biofunctionality. Indeed DNA
solitary waves (solitons), and in particular, the nucleotide
waves of oscillatory rotation, "read" the genome's sign
patterns, so that such sign vibratory dynamics may be considered
as one of many genomic non-linear dynamic semiotic processes.
The expression "DNA's texts";, borrowed earlier as a metaphor
from the linguists, is it turns out therefore related directly
to actual human speech. For as mathematical-linguistic research
into DNA and human speech textual patterns, shows [Maslow,
Gariaev 1994] the key parameter of both such patterns is
fractality. It can therefore be hypothesized that the grammar of
genetic texts is a special case of the general grammar of all
human languages.
Returning
however to DNA computation based on matter-wave sign functions
with a view to realizing its wave coding capabilities, as
distinct those used by Adleman, which might be termed its matter
capabilities. Such true wave control capabilities of the DNA or
chromosomes are, we hypothesize, those conditions that apply
inside the living cell, i.e. in an aqueous solution but which
correspond to a liquid-crystal condition as well. For under such
conditions, in the unique circumstances of cell division, the
living cell has the ability to replicate itself, and has the
property of what in relation to a self replicating automaton,
von Neumann [1966] called "universal computer construction" so
that we may say that the living cell is such a computer based on
DNA [Marcer Schempp 1997a]. And while the artificial cloning of
a single cell is not yet feasible, what we have been able to do,
is to record the DNA-wave information appropriate to these wave
sign conditions of the DNA in a cell on laser mirrors, and to
use, for example, the recorded DNA-wave information from living
seeds in the form of radio waves to resuscitate the
corresponding "dead" seeds damaged by radioactivity.
The
next step forward is therefore to bring into general use, such
wave information and memory as now newly identified in relation
to DNA and gene structure. Such applications could be on the
basis of, for example,
i)
The FPU-recurrence phenomenon, and/or,
ii)
The ability to record holograms, as well as,
iii)
The recording the polarization-wave DNA's information onto
localized photons.
Regarding
volume and speed, such memory could exceed many times over the
now available magnetic and optical disks, as well as current
classical holographic systems. But in particular, such
applications may employ the principles of quantum nonlocality.
For DNA and the genome have now been identified as active
"laser-like" environments, where, as experimentally shown,
chromosome preparations may act as a memory and as "lasers",
with the abilities i), ii) and iii) above. And finally there are
the quasi-speech features of the DNA, as these concern both
natural gene texts, and artificial (synthesized) sign sequences
of polynucleotides, which emulate natural quasi-speech gene
programs. However, we believe this maybe a rather dangerous
path, where a regulatory system of prohibitions on artificial
wave genes is indispensable. The reason is that such an approach
to DNA-wave biocomputation means entering new semiotic areas of
the human genome and the biosphere in general; areas, which are
used by the Nature to create humankind. This thought follows
from the theoretical studies on a collective symmetry of the
genetic code as carried out by the Eigen's laboratory [Scherbak,
1988] at the Max Planck Institute in Germany. This research
shows, that the key part of the information, already recorded
and still being recorded as quasi-speech in the chromosomes of
all organisms on our planet, may concern semantic exobiological
influences, since in regard to DNA-wave biocomputation, DNA acts
as a kind of aerial open to the reception of not only the
internal influences and changes within the organism but to those
outside it as well. Indeed we regard this as one of our primary
findings, which in view of quantum nonlocality of organisms
extends not only to the organism's local environment, but also
beyond it to the extent of the entire universe.
With
reference to what we have said already, it is possible to offer
the following perspectives on the sign manipulations with gene
structures.
1.Creation
of artificial memory on genetic molecules, which will indeed
possess both fantastic volume and speed.
2.Creation
of biocomputers, based on these totally new principles of
DNA-wave biocomputation, which use quantum teleportation
[Sudbury 1997] and can be compared to the human brain regarding
methods of data processing and functional capabilities.
3.The
implementation of a remote monitoring of key information
processes inside biosystems by means of such artificial
biocomputers, resulting in treatments for cancer, AIDS, genetic
deformities, control over socio-genetic processes and eventually
prolongation of the human life time.
4.Active
protection against destructive wave effects, thanks to
wave-information channel detectors.
5.Establishing
exobiological contacts.
2. What Experiment Confirms, part two, the Experiments
Some
of the experiments and computer simulations carried out in
Moscow are now described. They set out in more detail how the
understanding in sections 1. was arrived at. These descriptions
concern the specific apparatus used and results obtained,
together with computer simulations carried out to validate
specific aspects of the developing understanding,
Photograph
1. This first picture shows a photograph of the experimental
apparatus. The principal elements are a laser, the light of
which is directed through a lens system and a DNA sandwich
sample as shown diagrammatically below
Diagram 1. Illustrates the
workings of the experiment which employs a dynamic light
scattering system of the type Malvern.
This
understanding is then compared in section 3 with an entirely
independently researched prospective obtained by Marcer, and
Schempp [1996].
This
shows the scattering by the DNA sample of the laser light, which
is then guided through another lens system into the type Malvern
analysing device, which counts the photons registered in
different serial channels.The results of two experiments are
shown at end of paper: the first entitled "Background - Empty
Space", done without a DNA sample, and the second, with it in
place, entitled "Physical DNA in SSC Solution".
The
latter has the typical form of a periodically reoccurring
pattern, which is of the same functional type as found in an
autocorrelation. Such regularly occurring periodic patterns have
an interpretation in terms of the phenomenon of so-called
Fermi-Pasta-Ulam recurrence, which concerns solitonic waves.
That is to say, this interpretation says that roughly speaking,
the DNA, considered as a liquid-crystal gel-like state, acts on
the incoming light in the manner of a solitonic Fermi-Pasta-Ulam
lattice, as illustrated here:
***
The
leading question, if this is the case, is what could such action
achieve? The starting idea was that it must be concerned with
the reading of the genetic texts encoded in the DNA, where
however this language metaphor is now applied directly to these
texts. That is to say, rather than the usual analogy taking such
texts as a digital computer language or symbolic instruction
code, such texts are considered instead as having the semantic
and generative grammatical features of a spoken or written
context dependent human language. That is, we conceived of the
DNA acting in the same way as the human would, when presented
with a text from a good book on a fascinating theme, which, as
it is read, invokes actual 3 dimensional pictures/images in the
mind's eye.
The
reason for this choice concerned the problem in DNA coding
raised by the question of synonymy and homonymy as it applies to
the third element/codon of the codon triplets. For while, see
figure below, synonymy even seems to provide a kind of
redundancy, homonymy constitutes a serious difficulty under the
often proposed postulate that only the first two elements of the
DNA codon triplet (standing for a particular protein- the
picture in the mind's eye, so to speak) are the significant
ones. That is to say, how does the reading ribosome know which
protein has to be generated, if the third nucleotide in codon's
triplet does not of itself provide the answer with total
certainty? The proposed answer was, that this ambiguity might be
resolved by some kind of context dependent reading similar to
that inherent in human speech and language understanding.
Figure: Synonymy versus Homonymy
Satisfyingly,
this need to explain how such context-dependent reading might be
implemented in the DNA reduplication/reading process, as will be
shown, led back to the experimental evidence as presented above,
for it supports the postulate that such context dependent
reading of the DNA is indeed best understood in the framework of
a biosolitonic process model.
A soliton is an ultra
stable wave train often with a seemly simple closed shape, which
can arise in the context of non-linear wave oscillations. It
actually consists of a rather complexly interrelated assembly of
sub wave structures, which keep the whole solitonic process in a
stationary state over a comparatively long time. In the
literature, a soliton is often described as an entity, which is
neither a particle nor a wave in much the same way as is a
quantum, for it, too has wave/particle duality. It can also be a
means to carry information. Solitonic processing in DNA, would
therefore, it was hypothesized, relate, in one of its aspects,
the reading of the codons, to quantum computing [Patel 2000],
and this could therefore concern the soliton viewed as the
travelling "window", that opens in the double helix structure as
the reading takes place, as is illustrated below:
It
was therefore decided to model this reading process as a complex
mechanical oscillator [Gariaev 1994], capable of producing
solitonic wave transmissions, which takes the form of a system
of rotary pendulums, like those in a certain type of pendulum
clock, as illustrated,
to see if the computer
simulations could shed more light on just what might be
happening in the DNA. In the basic model, illustrated and shown
below, each of the oscillatory movements of each element of the
linked chain of oscillators depends heavily on the motion of its
neighbours, and on the differences in the specific weights of
the elements. Imagine now that the DNA forms such a kind of
pendulum, whilst the intertwined helices/chains are opened at
one particular section to provide the travelling window, as in
the previous figure. That is to say, the model to be simulated
is a chain of non-linear oscillators, the four types of which
can be identified with the Adenine (A), Cytosine (C), Guanine
(G), and Thymine (T) or Uracil (C) components DNA, all having
different spatial structures and masses, and where there is a
travelling window opened in the double helix. Such a model
allows a rather complex pattern of oscillation in the DNA chain
of elements, depending on the actual layout of the elements as
specified by the actual genetic code sequence involved. The
window as it travels, is therefore highly context dependent.
Starting
at the following sequence:
|
(5' начало) GGC CTA TGT GGA GAG GAT GAA CTA CGT GCA CCG AGA CCT GCG GGC GGC CAA CAT CCT GGT GGG GGA GAA CCT GGT GTG CAA GGT GGC TGA CTT TGG GCT GGC ACG CCT CAT CGA GGA CAA CGA GTA CAC AGC ACG GCA AGG TGC AAG TTC CCC ATC AAG TGG AGA GCC CCC GAG GCA GCC CTC TAT GGC CGG TTC ACC ATC AAG TCG GAT GTC TGG TCC TTC GGC ATC CTG CTG ACT GAG CTG ACC ACC AAG GGC CGG GTG CCA TAC CCA GGG ATG GGC AAC GGG GAG GTG CTG GAC CGG GTG GAG AGG GGC TAC CGC ATG CCC TGC CCG CCC GAG TGC CCC GAG TCG CTG CAT GAC CTT ATG TGC CAG TGC TGG CGG AGG GAC CCT GGA GGA GCG GCC CAC TTT TCG AGC TAC CTG CAG GCC CAG CTG CTC CCT GCT TGT GTG TTG GAG GTC GCT GAG TAG TGC GCG AGT AAA ATT TAA GCT ACA ACA AGG CAA GGC TTG ACC GAC AAT TGC ATG AAG AAT CTG CTT AGG GTT AGG CGT TTT GCG CTG CTT CGC GAT GTA CGG GCC AGA TAT ACG CGT ATC TGA GGG GAC TAG GGT GTG TTT AGG CGA AAA GCG GGG CTT CGG TTG TAC GCG GTT AGG AGT CCC CTC AGG ATA TAG TAG TTT CGC TTT TGC ATA GGG AGG GGG AAA TGT AGT CTT ATG CAA TAC TCT TGT AGT CTT GCA ACA TGG TAA CGA TGA GTT AGC AAC ATA CCT TAC AAG GAG AGA AAA AGC ACC GTG CAT GCC GAT TGG TGG AAG TAA GGT GTA CGA TCG TGC CTT ATT AGG AAG GCA ACA GAC CGG GTC TGA CAT GGA TTG GAC GAA CCA CTG AAT TCC GCA TCG CAG AGA TAT TGT ATT TAA GTG CCT AGC TCG ATA CAA TAA ACG CCA TTT GAC CAT TCA CCA CAT TGG TGT GCA CCT GGG TTG ATG GCT GGA CCG TCG ATT CCC TAA CGA TTG CGA ACA CCT GAA TGA AGC AGA AGG CTT CAT --- 1020 (3'-конец) |
the
figures, which follow, are those of the computer simulation of
this process of the travelling window, carried out in relation
to a particular fragment of viral DNA. The first two figures
with respect to the simulation, where the vertical is the time
axis, show what would happen, in case of a context dependent
reading beginning from two different nucleotides of the DNA
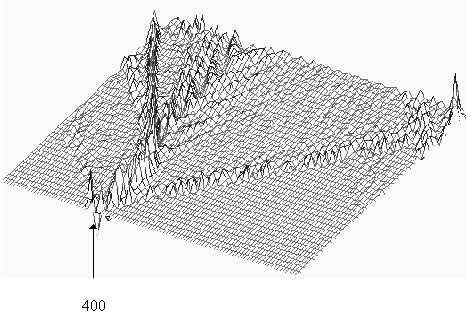
chain, namely the 400th and the 450th respectively. In both
cases these concern activity in the form of a "kink", which runs
through the chain of nucleotides, A, C, G, T. The second two
figures show even more sophisticated types of context dependent
effects. These concern the complex dynamic patterns, which arise
when also taking into account the non-linear covalent
connections between the nucleotides.

i) The masses of the nucleotides and other
parameters show that these oscillatory activities should be
located somewhere together in the "acoustic" wave domain, and
ii) That, as a liquid crystal, the DNA could
influence the polarization of the weak light emission known to
exist in cells, the so called "biophotons". This kind of emitted
light in cells was first discovered by the Russian investigator
Alexander Gurwitsch [1923], who called it the "mitogenic
radiation". Today it is known from the work of Fritz Albert Popp
[Popp, 2000], that such biophotonic or mitogenic light, while
being ultraweak, is however on the other hand, highly coherent,
so that it has an inherent laser-like light quality.
The
experimental setting and the resulting simulations therefore say
that:-
iii)
The experimental laser beam is simply a substitute for the
endogenous intracellular coherent light emitted by the DNA
molecule itself, and that
iv)
The superimposed coherent waves of different types in the cells
are interacting to form diffraction patterns, firstly in the
"acoustic" domain, and secondly in the electromagnetic domain.
Furthermore such diffraction patterns are by definition (and as
is known for example from magnetic resonance imaging (MRI)
[Binz, Schempp 2000a,b] a kind of quantum hologram. Thus, it
seems that our original picture is confirmed and that the
considered interaction between solitonic oscillations in the
liquid crystal structure of DNA, and the polarization vector of
the ultraweak biophotonic highly coherent light, could indeed be
hypothetically understood as a mechanism of translation between
holograms in the "acoustic" frequency domain, which concerns
rather short range effects and those in the electromagnetic
domain and vice versa.
The
basis of such an hypothetical mechanism as a translation
process, between acoustic and optical holograms, can be easily
illustrated in the laboratory, where, as shown below, there is a
fish illuminated in water by means of the acoustic radiation, in
such a way that on the surface of the water an interference
pattern or hologram forms, such that when this interference
pattern is illuminated from above in the right way, by light of
a high laser quality, a virtual visual image of the fish appears
above the water. It shows that the hologram in question acts as
a holographic transducer between the acoustic and
electromagnetic domains.

Laboratory illustration of a holographic transducer between the acoustic and electromagnetic domains
This illustrated transduction when described in terms of the formalization of Huygens' principle of secondary sources [Jessel 1954], has been used as the basis of a new topological computing principle [Fatmi, Resconi 1988] which defines entire classes of non-commutative control structures, Fatmi et al [1990]. It was applied to DNA. and more recently to the brain [Clement et al. 1999].
3.
Another Theoretical but Experimentally Validated Perspective -
Quantum Holography
Sections
1 and 2 are in excellent agreement with the independently
researched model of DNA produced by Marcer and Schempp [1996].
This explains the workings of the DNA-wave biocomputer in terms
of a quantum mechanical theory called quantum holography
[Schempp 1992] used by Schempp [1998] and Binz and
Schempp [2000a,b; 1999] to correctly predict the workings of
MRI. These two DNA-wave biocomputer models are also, as cited,
in good agreement with qubit model explanation of DNA more
recently published by Patel [2000], and earlier independent
researched models by Clement et al [1993] and Perez [1991].
The quantum holographic
DNA-wave biocomputer model describes the morphology and dynamics
of DNA, as a self-calibrating antenna working by phase conjugate
adaptive resonance capable of both receiving and transmitting
quantum holographic information stored in the form of
diffraction patterns (which in MRI can be shown to be quantum
holograms). The model describes how during the development
of the embryo of the DNA's organism, these holographic patterns
carry the essential holographic information necessary for that
development. This would explain the almost miraculous way the
multiplying assembly of individual cells is coordinated across
the entire organism throughout every stage of its development -
in complete agreement with the explanation arrived at in Moscow
by Gariaev and his co-workers
The quantum holographic
theory requires that the DNA consists of two antiparallel
(phase conjugate) helices, between which (in conformity with
DNA's known structure, ie the planes on which the base pairing
takes place) the theory says, are located hologram
planes/holographic gratings, where the necessary 3 spatial
dimensional holographic image data of the organism is stored in
agreement with the Gariaev group's hypothesis. It says, as
described in relation to laser illumination of a DNA sample,
that such illumination can be expected to turn the DNA into a
series of active adaptive phase conjugate mirrors (see figure
below)/holographic transducers (see figure of laboratory
illustration earlier), from which would resonantly emerge a beam
of radiation, on which is carried the holographic information as
encoded in the DNA. As indeed is the case in the Gariaev group
experiments already described. These experiments thus confirm
the quantum holographic prediction that DNA functions an
antenna capable of both encoding and decoding holographic
information. This functionality is also in good agreement with
the findings of Schempp [1986] that quantum holography
is capable of modelling antennae such as synthetic aperture
radars, and that this mathematical description of radar can be
applied [Marcer and Schempp 1997] to a model, working by quantum
holography, of the neuron. This model is in good accord
with the biological neuron's information processing morphology
and signal dynamics. As indeed are the quantum holographic
models of the brain as a conscious system, and of the prokaryote
cell [Marcer, Schempp 1996, 1997a]. It is a viewpoint originally
voiced by de Broglie, who presciently pictured the electron as
being guided by its own pilot wave or radar! These examples
including MRI all demonstrate that quantum holograph does
indeed incorporate signal theory into quantum physics and it can
be hypothesized biocomputation.
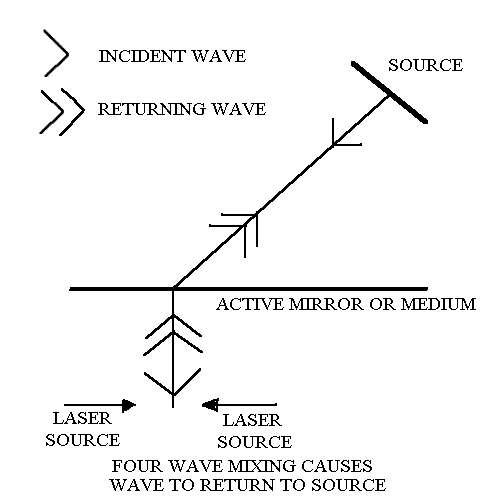
Phase conjugate mechanism or mirror in the laboratory. Action of an active adaptive phase conjugate mirror
Furthermore, quantum holography predicts that the planes, in which the base pairing takes place, constitute a "paged" associative holographic memory and filter bank (carrying holograms which can be written and read) and which has no cross talk between the pages. The orthogonality of the holograms encoded on these pages, arises as the result of the sharp frequency adaptive coupling conditions (1), which specify very narrow spectral windows, i.e. the "pages".
(1) <Hv(a,b; x,y)| Hv(c,d ; x,y)> = 0 when frequency v is not equal v'
<Hv(a,b;
x,y)| Hv(c,d ; x,y)> = <aOb | cOd> when v = v'
for
non-degenerate four wavelet mixing where a,b,c,d are the
corresponding wave functions of the mixing; Hv(a,b; x,y) is the
holographic transform which in quantum holography defines
the probability of detecting a wave quantum frequency v within a
unit area attached to the point (x,y) of the hologram plane,
where the wavelet mixing aOb takes place and is described in
terms of a tensor multiplication O. The orthogonality condition
(1) can be seen therefore as specifying a set of diagonal
elements or trace Tr in a unit matrix in the frequency domain.
It implies, as can be shown, that the Shannon encoding schema
employed in DNA is optimally efficient, which following a
billion or more years of evolution, in DNA could be expected to
be the case.
The
conditions (1) are therefore in excellent agreement with Gariaev
group's conclusion. It confirms that the planes on which the
base pairing takes places, concerns two quantum holograms, ie
the wavelet mixings aOb and cOd, where each specifies a
"context", one for the other. Further quantum holography predicts,
based on the symmetries of the 3 dimensional representation of
the Heisenberg Lie group G, that in relation to the quantum
hologram defined by a wavelet mixing aOb, the coherent wavelet
packet densities a(t)dt and b(t')dt' are indistinguishable by
means of relative time and phase corrections applied to the
respective wavelet pathways (x,y) in the hologram plane. That
is, to say, the tensor operation O, in the case of quantum
holography, describes a quantum entanglement, even though
aOb defines a quantum hologram, from which quantum
holography shows and MRI proves, holographic information
can be both written/encoded and read/decoded.
Thus,
mathematically, DNA can on the basis of quantum holography
be thought of represented quantum mechanically very simply by
the trace
Tr < a,b | c,d >
such
that when the double helix is opened, in accordance with the
Gariaev description above, this corresponds to the
representation
< a,b | >< | c,d >
The process of completed duplication of DNA can
therefore represented as
Tr<a,b
| c,d>< a,b | c,d >
The
topological differentiation referred to above follows from the
fact that, while in quantum mechanics, a wave function is only
determined up to an arbitrary phase, phase difference is of
physical significance (as in holography), because there exists a
class of quantum observables, which are the gauge invariant
geometric phases of the state vector or wave function [Resta
1997; Schempp 1992; Anandan 1992]. These observables must
therefore be distinguished from those which are the eigenvalues
of some operator, usually the Hamiltonian or energy function.
Such a state vector description (with gauge invariant phases) by
means of which each DNA molecule can clearly be expected to be
described, would explain the difference between the nature of
quantum interference and quantum self interference, which DNA
from its double helical structure can thus be recognized to
concern.
In
the above means of representing DNA therefore, | >< |
represents by the quantum correspondence principle, the quantum
soliton control [see also, Denschlag et al, 2000] or wavepacket
activity rather than its classical soliton counterpart, which
was the subject of the Moscow computer simulations. These all
confirm the Gariaev group's conclusions reached as a result of
their experiments, that DNA functions as a quantum coherent
system/assembly (of now quantum oscillators) or whole, by means
of quantum entanglement. A whole, where as (1) shows, this may
be decomposed into an orthogonal family of holographically
encoded 3 spatial dimensional images in line with the usual
description of a quantum mechanical diagonalization. It also
says in line with the Gariaev group's findings that DNA can be
described as an "autocorrelation", where as shown here, this is
an optimally efficient decomposition into a decorrelated family
of holographic code primitives /holograms, and that this, as
Schempp[1992] shows, follows from the fact a quantum mechanical
harmonic oscillator (in this case the highly complex DNA
molecule itself) is equivalent to an assembly of bosons each
having one polarization state. The latter substantiates the
Gariaev group conclusion that they have indeed discovered an
entirely new form of electromagnetic vector by means of which
holographic images are carried in the form of a polarization
state, suitable for a new form of cinema, video and computer.
Quantum
holography says that DNA satisfies the principle of
computer construction [Von Neumann, 1966], since it carries a
copy of itself, and is
Graphs
(a), (b), and (c) : 'Background – Empty Space', 'Physical DNA
in SSC Solution', and 'Phantom DNA' respectively

Graphs (a), (b), and (c) Background – Empty Space, Physical DNA in SSC Solution, and Phantom DNA” respectively


References
Adleman L.M. 1994, Molecular Computation of
Solutions to Combinatorial Problems, Science, 266, 11th November
1021-1024.
Allison S.A., Sorlie S., Pecora R., 1990,
Macromolecules, v.23, 1110-1118.
Anandan J. 1992, The geometric phase,
Nature, 360, 26, 307-313
Binz E. Schempp W. 2000a Creating Magnetic
Resonance Images, Proceedings CASYS'99, International Journal of
Computing Anticipatory Systems, 7, 223-232.
Binz E. Schempp W. 2000b, A Unitary
Parallel Filter Bank Approach to Magnetic Resonance Tomography,
American Institute of Physics Proceedings 517 of the 3rd
International Conference on Computing Anticipatory System,
editor Dubois D. August 9-14, 1999, Liege, Belgium, 406-416.
Binz E., Schempp W. 1999, Quantum
Teleportation and Spin Echo, Unitary Symplectic Spinor Approach.
In. Aspects of Complex Analysis, Differential Geometry,
Mathematical Physics and Applications, Dimiev S. Sekigawa K.
editors, World Scientific, 314-365.
Bouwmeester D. et al., 1997, Experimental
Quantum Teleportation, Nature, 390, 11th December 575-579
Clement B.E.P. Coveney P.V. Marcer P. 1993,
Surreal numbers and optimal encodings for universal computation
as a physical process: an interpretation of the genetic code.
CCAI Journal, 10, 1/2, 149-164.
Clement B.E.P. Coveney P.V, Jessel M. and
Marcer P. 1999, The Brain as a Huygens' Machine. Informatica 23,
389-398.
Denschlag J. et al, 2000, Generating
Solitons by Phase Engineering of a Bose-Einstein Condensate,
Science, 287, 7th January, 97-101
Dubois D., 1992, The Fractal Machine, Liege
University Press, Liege.
de Duve, C. 1984 A Guide Tour of the Living
Cell, volume two, Scientific American Library,
Fatmi H.A. and
Resconi G. 1988, A New Computing Principle, Il Nuovo Cimento,
101B, 2, 239-242.
Fatmi H.A., Jessel
M., Marcer P.and Resconi G. 1990, Theory of Cybernetic and
Intelligent Machine based on Lie Commutator. International
Journal of General Systems, 16, 123-164.
Fermi E., 1972,
Proceedings. Moscow. Science. v.11.
Gariaev P.P.,
Junin A.M., 1989, Energy, no10,
46-52. [in Russian]
Gariaev P.P.,
Chudin V.I., Komissarov G.G., Berezin A.A., Vasiliev A.A., 1991,
Proc. SPIE, v.1621, 280-291.
Gariaev P.P, 1994,
Wave genome, Public Profit. Moscow. 279 pages [in Russian].
Gurwitsch A.,
1923, Versuch einer synthetishen Biologie. Schaxels Abh. Z.
theor. Biol. H. 17.
Ho M-W, 1993, The
Rainbow and the Worm: The Physics of Organisms, World
Scientific, Singapore.
Jessel M. 1954, Une formulation analytique
du principe de Huygens, Comptes Rendus, 239, 1599-1601.
Marcer P. 1992, Designing New Intelligent
Machines - The Huygens' Machine, CCAI
Journal for the Integrated Study of
Artificial Intelligence and Applied Epistemology, 9,4, 373-392.
4, 373-392.
Marcer P. 2000, Hypercomputation, Third
International Conference on Computing Anticipatory Systems,
CASYS '99, Liege, Symposium 9 on Quantum Neural Information
Processing, Journal of Computing Anticipatory Systems, ed.
Dubois D., vol. 7, 288-312.
Marcer P. this volume, Quantum Millennium,
Quantum Universe, Quantum Biosphere, Quantum Man.
Marcer P. (in press) Anticipation and
Meaning, CASYS 2000, International Conference on Computing
Anticipatory Systems, AIP Conference Proceedings, ed Dubois D.
Marcer P. and Schempp W., 1996, A
Mathematically Specified Template For DNA And The Genetic Code,
In Terms Of The Physically Realizable Processes Of Quantum
Holography, Proceedings of the Greenwich Symposium on Living
Computers, editors Fedorec A. and Marcer P., 45-62.
Marcer P., Schempp W. 1997, Model of the
Neuron working by Quantum Holography, Informatica 21, 5 19-534.
Marcer P and Schempp W., 1997a, The Model
of the Prokaryote cell as an Anticipatory System Working by
Quantum Holography, Proceedings of CASYS 97, 11-15, August,
HEC-Liege, Belgium, International Journal of Computing
Anticipatory Systems, v.2, 307-315.
Marcer P., Schempp W. 1998, The brain as a
conscious system, International Journal of General Systems, 27,
1/3, 231-248.
Maslow M.U., Gariaev P.P., Fractal
Presentation of Natural Language Texts and Genetic Code, 2nd
International Conference on Quantitative Linguistics", QUALICO
'94, Moscow, September 20-24, 193-194, 1994
Patel A. 2000, Quantum Algorithms and the
Genetic Code, Proceedings of the Winter Institute of Quantum
Theory and Quantum Optics, 1-13 January, S.N. Bose National
Centre for Basic Sciences, Calcutta, India.
Perez J-C, 1991, De l'ordre et du chaos
dans l'ADN, Science et Technology, April, 36, 40-47.
Pribram K.H. 1991, Brain and Perception;
Holonomy and Structure in Figural Processing, Lawrence Eribaum
Associates, New Jersey.
Popp F.A., 2000, Some features of
biophotons and their interpretation in terms of coherent states.
Biophotonics and Coherent Systems. Proc. 2nd A.Gurwitsch
Conference and Additional Contributions. Moscow University
Press. Ed. L.Beloussov et al., 117-133.
Resta R., 1997,
Polarization as a Berry Phase,(The Berry Phase), Europhysics
News, 28,19
Rice S.A. 1992,
New Ideas for Guiding the Evolution of a Quantum System,
Science, 258, 16th October, 412-413.
Schempp W. 1986,
Harmonic Analysis on the Heisenberg Group with Applications in
Signal Theory, Pitman Notes in Mathematics Series, 14, Longman
Scientific and Technical, London.
Schempp W. 1992,
Quantum holography and Neurocomputer Architectures, Journal of
Mathematical Imaging and Vision, 2, 279-326.
Schempp W., 1993,
Bohr's Indeterminacy Principle In Quantum Holography,
Self-adaptive Neural Network Architectures, Cortical
Self-organization, Molecular Computers, Magnetic Resonance
Imaging and Solitonic Nanotechnology, Nanobiology 2, 109-164.
Schempp W. 1998,
Magnetic Resonance Imaging, Mathematical Foundations and
Applications, John Wiley, New York.
Scherbak V.I.,
1988, J. Theor. Biol., v.132, 121-124.
Schleich W.P.
1999, Sculpting a Wavepacket, Nature, 397, 21st January,
207-208.
Sudbery T. 1997,
The Fastest Way from A to B., Nature, 390, 11th December,551-552
Sutherland J. 1999, Holographic/Quantum Neural Technology, Systems and Applications, an ANDCorporation documented plenary presentation for the 3rd International Conference on Computing Anticipatory Systems, 8-14 August, HEC Liege, Belgium. International Journal of Computing Anticipatory System, 2000, 7, 313-334, sutherland@andcorporation.com. and http://www.andcorporation.com.
Von Neumann J. 1966, Theory of A Self-reproducing Automaton, University of Illinois Press, Urbana and London.
Vladimir POPONIN Patents
US2002123050
Apparatus and method for analysis of nucleic acids hybridization on high density NA chips
Inventor: POPONIN VLADIMIR
2002-09-05
RU2000127
METHOD OF CONTROLLING FUNCTIONAL CONDITION OF BIOLOGICAL OBJECT MAINLY FOR CORRECTION OF PATHOLOGICAL CONDITION
RU2000126
SYSTEM OF CONTROLLING FUNCTIONAL CONDITION OF BIOLOGICAL OBJECT MAINLY FOR CORRECTION OF PATHOLOGICAL CONDITION
RU2005509
METHOD FOR CONTROL OF FUNCTIONAL CONDITION OF BIOLOGICAL OBJECT PREDOMINANTLY FOR CORRECTION OF PATHOLOGY
RU2004266
METHOD OF CONTROLLING FUNCTIONAL CONDITION OF BIOLOGICAL OBJECT FOR CORRECTION OF PATHOLOGIC CONDITION
The Wave, Probabilistic and Linguistic Representations of Cancer and HIV
by
Peter P. Gariaev*, George G. Tertishny, Katherine A. Leonova
*Chief Scientific Officer,
Wave Genetics Inc.,
907 Alness Street, North York, ON M3J 2N2, Toronto, Canada.
(416) 6616614
Abstract: The basic assumptions of
our work include the following: 1. the genome has a capacity
for quasi-consciousness so that DNA “words” produce and help
in the recognition of “semantically meaningful phrases”; 2.
the DNA of chromosomes control fundamental programs of life in
a dual way: as chemical matrixes and as a source of wave
function and holographic memory; 3. processes in the
substance-wave structures of the genome can be observed and
registered through the dispersion and absorption of a bipolar
laser beam. The present article brings forward considerable
theoretical and experimental evidence in support of this
model, and discusses its practical applications with respect
to cancer and HIV therapeutic strategies.
I. The nature of HIV and
cancer: problems in interpretation.
The challenge of HIV and cancer and the essence of Life both lie on the same plane. As of now, we still don't understand the most crucial facts about Life: how did it appear on earth and in which way it is coded in chromosomes? Several hypotheses are available, and each of them at best represents just a piece of reality. This is where the theoretical and biological difficulties in interpreting the HIV and cancer phenomena come from - and the price we are paying for this misunderstanding are the mistakes emerging in the treatment of these diseases. Both pathologies affect the most vital part in any biological system, namely, its genetic apparatus - the organism’s “self-knowledge”. And this is the paradox: we seem to know quite enough about chromosomes and DNA - oncogenes have already been found, the HIV genome has been studied, and it’s clear how these informational structures function in chromosomes. The genetic code and ribosome operation principles also seem to have been investigated in detail. But for some reason all this information is not enough to develop universal methods for a successful intervention against cancer and HIV.
A half-lie is the worst lie, because people have reason to believe it - and this is especially true with respect to the genetic coding paradigm. In this field, everything is an impregnable bastion for critics, and everything is ruled by dogma. Even the key definition, the strategic scheme of genetic coding (DNA-> RNA-> protein), is called “the Central Dogma”. Until recently, all attacks on this dogma seemed trivial and doomed to failure. That was an incorrect assumption, as it turned out. The accuracy and effectiveness of research strategies dealing with HIV, cancer and many other pathologies depend on whether we orderly understand the genetic coding mechanism. The discovery of reverse transcriptase was the first spectacular breach in this dogma, which as a result was re-assigned a more discreet, working-hypothesis status: DNA Û RNA--> protein. However, our ideas on protein biosynthesis are gradually eroding: each new model is just an approach to the truth, to the understanding of the genome language-image pluralism as coding tool for the spatiotemporal structure of biosystems [32, 33].II. What do we want to prove?
In this paper we intend to propose ideas which are not aimed at the final destruction of the so-called genetic code “canonical” triplet model, but at the development and establishment of its exact position in the knowledge basis of the chromosome operational principles. Yes, it’s possible to state that the triplet code is the truth. However, this truth is as correct as the statement that we could write a word using an alphabet. That statement is certainly right. But if we try, based only on this knowledge, to go further and vouch that by means of this alphabet we can compile grammatically-correct sentences, this new statement won’t be accurate. Such a statement is incorrect, in fact, because for the compilation of human speech laws of syntax, logic and grammar have to be applied. As for the genome, it’s a very speech-like and logical structure, but its fundamental features are not the only way to express genome associative-semantic structures. Furthermore, we are inclined to agree with V.V.Nalimov’s ideas [43] leading us to the conclusion that a genome possesses quasi-conscious abilities. The logic we use and the models we developed are only an attempt to obtain higher-level understanding of laws pertaining to genetic text structuring or to other genome vital structures - knowledge which is now just beginning to emerge. The Russian researchers A.G.Gurwitch [38], V.N.Beklemishev [29] and A.A.Lyubitchev [41] laid the foundation of this science in the late 1920s.
What type of approaches might enrich the commonly-accepted genetic coding theory and how can these innovations assist in resolving the HIV and cancer issues, in particular? Let’s assume, until getting a final proof, three "axioms" which have already gotten definite theoretical and experimental confirmation [8, 32, 33, 37]:
DNA molecules, included in chromosomes, possess a substance--wave duality which is similar to the dualism of elementary particles. In accordance with it, DNA codes an organism in two ways, both with the assistance of DNA matter and by DNA sign wave functions, including coding at its own laser radiation level - [28].
The genetic apparatus can be non-local at the molecular level (holographic memory of a chromosome continuum) and at the same time quantum mechanically non-local in compliance with the Einstein-Podolsky-Rosen effect [4]. The latter means that the genome genetic and other regulatory wave information is recorded at the polarization level of its photons and is non-locally (everywhere and in no time) transferred (plays out) throughout the entire space of a biosystem by the polarization code parameter. This helps to set a quick-response information contact among the billions of cells constituting an organism.
The genome on the whole and the individual nucleus
of cells can generate and recognize text-associative regulatory
structures with the application of a background principle,
holography and quantum non-locality.
III. What’s the next step?
Let’s assume that final proofs of the above-mentioned statements have been obtained. Then the problem of HIV and cancer rises to an altogether different intellectual dimension.
For instance, what does the “DNA matter-wave dualism” mean - and in which way is it linked with the chromosome's numerous code functions? (Note: by code function we understand processes which are dramatically differed from the known triplet genetic code) In some sense, the genome operates like a complex multiwave laser with adjustable frequencies. It emits DNA light which is gene- and sign-modulated by amplitude, phase, frequency and polarization. Moreover, the genome is also likely to be a radio wave emitter converting a wide spectrum of coherent sign-polarized radio bands [37] (P.P. Gariaev, G.G.Tertyshniy, Ye.A. Leonova, etc. Radio wave spectroscopy of local photons: exit to quantum non-local bioinformational processes. Sensors and Systems (2000, N9, pp. 2-13). The genome is also a dynamical multiple hologram which is able to produce light and radio wave images [37] which carry out management functions by the biosystem. These structures are also the carriers of electromagnetic marking schemes (calibration fields) of biosystems’ space and time organization. And finally, the genome is a quasi-text form possessing elements of quantum non-locality, which can without any time delay “read” itself in billions of cells and use information, thereby received, as a control blueprint for living functions and structural organization [8, 37]. Many biologists and geneticists, let alone doctors, are likely to consider these new concepts of genome organization as extremely complicated. However, not all of them will: these ideas, whose seeds were first planted in Russia in the 1920s, have seen a dramatic and accelerating development over the last decade.
This clearly suggests that it’s necessary to modify our strategy in searching for HIV and cancer cures, as the traditional approaches to solving this issue increasingly resemble the wish to produce a good harvest, having planted an asphalt road. The new strategy has to be based on fundamental investigations of substance-wave duality and quasi-speech attributes of a higher system genome. Let’s stress once again that we consider a chromosome continuum as a sign laser & radio wave emitter [8, 33, 37], and direct experimental evidences allows us to think so. For instance, to demonstrate laser abilities of genetic structures, we showed that DNA and chromatin in vitro could be pumped as a laser-active medium for a consequent laser light generation [28].
If we accept these vital characteristics of a genome, then new specific issues arise: for example, does the sign character of chromosome laser & radio wave radiation change when a xenobiotic HIV genome inserts itself into it? And, at the same time, what happens to the radiation “semantics” during a transposition of oncogenes or any other mobile polynucleotide sequences as well as during B<--> Z and other conformational transitions of DNA in-vivo? Are these changes linked to an alteration of quasi- and real holographic programs, i.e. are new programs created and old ones changed, or are these programs erased, and so on? Does the radiation polarization parameter retain, in semiotic sense, its dynamic properties in the process of genome reorganization? Do all these changes influence ribosome operation? Further questions may arise. The answer to any of them can play a key role in interpreting the nature of HIV and cancer.
IV. Theoretical structures - more details
Let’s take another fundamental problem. Oncogene and HIV genomes, occupying certain positions in a 3D space of master cell chromosomes, do not express themselves as pathogenic factors until a certain time. In this sense, the behavior of HIV in the infected patient’s organism is unpredictable. HIV's latency period may vary from a week to 10 years. A specific mechanism of HIV-infection induction from the latent (sleeping) condition is thought to exist, but this mechanism is still misunderstood and, therefore, the opportunity to make HIV viruses permanently latent in the human organism is being missed. In this condition, the organism and the cells simply “don’t notice” them or even, as in the case of oncogenes, use them for their own benefit as a reproduction factor. Why does an organism adequately accept and contain them until a certain time X, and why they are semantically reborn, causing a management catastrophe in cell, after the X-time has come? Following our logic, it’s possible to think that both in the pathologic and normal state four factors are engaged, at least: genome “holography” and “linguistics”, genome background (context) self-organization, and its quantum non-locality.
In the course of evolution, biosystems have produced their own genetic “texts” and a biocomputing genome as a quasi- intelligent “subject” which “reads and understands” these texts at its level. The fact that natural human texts (it doesn’t matter what the language is) and genetic “texts” have similar mathematical & linguistic and entropy-statistical characteristics is extremely important for the genome elementary “intelligence”. This relates, in particular, to concepts such as the fractality of letters' occurrence frequency density distribution (in genetic “texts”, nucleotides execute function of letters) [21].
American researchers obtained another confirmation of the genome coding function linguistic interpretation [20]. Dealing with the “coding” and “non-coding” eukaryote DNA sequences [(in the framework of old concepts of a gene), they came to a conclusion which was similar with ours and which conflicted with the central dogma that meaningful functions are concentrated only in the protein-coding DNA sections. The researchers applied a statistical analysis method for studying natural and musical texts, known as Zipf-Mandelbrot’s law, as well as the known Shannon postulate of text information redundancy calculated as a text entropy (more information about text entropy and statistics of words distribution in texts is given in [1, 25, 27, 31]). As a result, they found that DNA “non-coding” areas (space, intronic and others) had more in common with natural languages than the “coding” ones. Taking this for granted, the authors inferred that “non-coding” sequences of genetic molecules were the basis for one or more biological languages. Furthermore, the authors developed a statistical algorithm for searching DNA coding sequences; the algorithm they developed demonstrated that protein-coding areas had significantly fewer long-distance correlations, compared with areas separating these sequences. The DNA-sequence distribution was so sophisticated that the methods the researchers applied stopped working satisfactorily at distances of over 10^3-10^2 base pairs. Zipf-Mandelbrot’s distribution for “words” occurrence frequency, where the number of nucleotides ranged from 3 to 8, demonstrated that natural language had more in common with the non-coding sequences, than with the coding ones. It’s worth noting that the authors considered the coding only as a record of amino acid sequence information. And that was a paradox which made them state that DNA non-coding areas were not merely “junk”, but the lingual structures designed for reaching some still unknown goals. Despite the discovery of hidden complexity in these non-coding areas, the authors didn’t understand the implications of the long-distance correlations characterizing these structures. (They illustrated the process based on a family of genomes of the myosin heavy chain and assigned it to the evolutionary transition from lower taxons to higher taxons). The data presented in [20] is in full compliance with the ideas we had independently put forward [32, 33]; according to our point of view, DNA non-coding sequences, or approximately 95-98% of a genome, are a strategic informational content of chromosomes. The said context has a substance-wave nature and, therefore, is multidimensional and functions as a holographic associative-image and semantic program of embryological origin, the semantic continuation and the logic end of any biosystem. Having intuitively understood that the old genetic coding model led to a dead-end, the authors [20] said a nostalgic good-bye to this now-obsolete paradigm, but didn’t propose anything to replace it.
V. Homonymous-synonymous ambiguities of genetic texts. What does an organism need them for?
Text homonymy and synonymy are the common fundamental semantic properties of natural and genetic texts. These features provide chromosomes, natural texts and speech with redundant and multivalent information and, thus, ensure some adaptive flexibility. Ambivalent genetic texts acquire their monosemantic meaning owing to a variation of DNA sequence position in genome space through their transpositions and/or a transposition of their environment. This resembles the situation with natural texts and speech, in which homonymous-synonymous ambiguities of a semantic field are eliminated by the context (this background principle is described in [44]). Homonymies of coding doublets are easily found in the traditional genetic code triplet model. The meaning of these homonymies is still misunderstood and isn’t taken into account, with some exceptions [33, 35]. The perplexing issue of mRNA codon homonymies emerged with the creation of the triplet model of amino acid coding in the process of protein biosynthesis. It immediately became a “time-bomb”, since the correct explanation of a biological (informational) meaning of these homonymies automatically leads to the necessity of significant correction or complete revision of the triplet model. How are codons homonymies produced? A set of different amino acids is coded in mRNA codons by similar doublets; the third nucleotides in codons can relocate chaotically: they are wobbling and may become any of the four canonical ones. As a result, they don’t correlate with the coding amino acids [3, 11]. That’s why semantic ambiguity appears regarding the ribosome’s choice of amino acid-carrying-tRNA anti-codons. For instance, each synonymous codon of the standard code of higher biosystems (AGT and AGC) codes for serine, while each synonymous AGA and AGG codon codes for arginine. Thus, the third nucleotides of mRNA codons in combination with a sign doublet don’t have exact amino acid correlates; at the same time, the first two sign codon nucleotides are similar with one another, yet code different amino acids - hence the ambiguity in selecting tRNA anti-codons. In other words, a ribosome may take serine or arginine tRNA with an equal probability ; such an outcome can initiate synthesis of abnormal proteins. In fact, these mistake don’t occur and the precision of the protein synthesis process is extremely high. These mistakes appear only in some metabolically abnormal situations (the presence of some antibiotics, a lack of amino acids, etc.). Usually a ribosome somehow correctly chooses the tRNA anti-codons out of the homonymous doublets.
We think that the correct choice out of doublet anti-codon-homonyms is realized through a resonant-wave or context (associative, holographic) and/or “background” mechanisms. Amino acid code homonymity can be overcome in the same way as they are in natural languages - by the placement of a homonym in a complete phrase; the homonym decodes the context and attaches a unique meaning to it, thus resolving the ambiguity. That’s why mRNA, acting as a “phrase”, should operate in the protein synthesis process as an integral coding system, non-locally determining the sequence of amino acids at the level of tRNA aminoacylated associates, which interact in a global and complementary way with the entire mRNA molecule. Macrosteric disagreement between mRNA and tRNA continuums could be eliminated due to a conformational lability of macromolecules. The A-P sections of a ribosome are responsible for accepting these associated amino acids, with their consequent enzymatic sewing into peptide chains. In this case, a context-oriented unambiguous choice and elimination of the doublet-anticodon homonymy will occur. Considering the above, it’s possible to predict that the interaction of aminoacylated tRNAs with mRNAs has a collective phase character and is effected by a type of re-association (“annealing”) of one-string DNA upon the temperature reduction after melting of a native polynucleotide. Does any experimental evidence for this contention exist? Yes. A great deal of such information is available and collected in the analytical review [45]. Here we will only present some of the data. For example, the correctness of terminating codons recognition by tRNA molecules is known to depend on their context (that’s a confirmation of our theoretical model), in particular, on the existence of a uridine after the stop codon. In Paper [9] the following information is presented: the insertion of a line consisting of nine rarely-used CUA-leucine codons in the position after the 13th one (in the compound of 313 codons of the tested mRNA) resulted in active inhibition of their translation, yet did not notably influence the translation of other CUA-codon-containing mRNAs. Here, the translation context effect is clearly seen as a strategic influence of distant mRNA codons on the inclusion (or non-inclusion) of certain amino acids in the composition of a protein being synthesized. This is a remote influence, connected with the protein synthesis continuum; it’s also an example of the genetic apparatus’ non-local functions, whereby the protein-synthesizing apparatus recognizes mRNA not only in parts (by nucleotides, locally), but in one piece (non-locally) as well. However, in the work being cited this key phenomenon is only stated and remains inexplicable to the researchers; and probably for this reason they don’t even discuss it. Similar results continue to appear in the literature at an increasing rate. In the work under discussion the authors refer to half a dozen analogous situations, whose explanation in the classical interpretation is rather difficult. This obviously points to inconsistencies in the genetic code triplet model. The model also fails to explain the existence of unusually swollen anticodons. When they are involved in protein synthesis, the number of base pairs in the ribosome A-site exceeds 3 [45]. This finding challenges the dogmatic postulate of code triplets . Furthermore, studies of tRNA-tRNA interactions on ribosomes are presented in [45]; they offer full confirmation of our model, in which we consider an amino-acid-loaded tRNA complex as the predecessor of a protein. In [45] an important idea, very close to ours, was put forward: the influence of the mRNA context on monosemantic incorporation of amino acids into a peptide chain reflects some basic, still unstudied, laws of genetic information coding in the protein synthesis process. It’s worth remembering that genetic information about protein synthesis occupies only some 1% of a chromosome's total volume. The remaining 98.5% of the whole contain programs of a significantly higher level.
VI. Prions: the last blow to the central dogma of molecular biology
As we can see, the previously-existing hypotheses regarding the genetic code and the operation of the protein-synthesis apparatus have been grossly simplified. The prion phenomenon is likely to be the last argument in favor of a final revision of the molecular biology central dogma.
Prions are low-weight molecular parasitic proteins (PrPsc) targeting the brains of animals (mad-cow disease) and human beings (Alzheimer’s, Kreutzfeld-Jacob’s syndrome, etc.). Virus-like strain-specificity is an inexplicable feature of prions. This strain-specificity is only attributed to microorganisms or viruses which have a genetic apparatus. And yet, it’s thought that prions don’t have a genome, since all efforts to find traces of DNA or RNA in them have failed. An acute contradiction, which once again discredits the molecular biology central dogma, arises: prions don’t have a genome, but genetic signs are present. Some scientists, unable to explain this phenomenon and trying to “save” the central dogma, nevertheless suppose that DNA or RNA traces are hidden in the prion molecule’s wrinkles [10]. However, investigations carried out in this field over decades and endorsed by the Nobel prize awarded to Stanley Prusiner in 1997, reliably demonstrated that prions had neither nucleic acids nor a genome [23]. How are we to resolve this contradiction? If we admit that the central dogma is correct, then this is impossible. Having rejected this dogma, we can imagine the following prion biogenesis scenario [34]: in this model, a “prion virtual genome”, i.e. a provisional genome "borrowed" from the master cells for a given time, is the chief player. To put it more exactly, this is a protein-synthesizing apparatus of master cells. Prions are likely to have retained the paleogenetic way as their way of reproduction; in some cases this breeding method enables prions not to use genes encoded in chromosomes, but to self-reproduce in another way, ignoring the central dogma of molecular biology and genetics. To synthesize prions, a cell has to address their genes: it’s a progressive, but, at the same time, organizationally and energetically difficult method. Prions can simplify this procedure.
We believe that PrPsc (Prion-Protein-scrapy) NH-group peptide bonds can react with the OH-groups of ribose remains of accepting CCA-sequences of respective tRNAs. In the course of a hypothetical fermentative reaction, an emerging poly-tRNA-complex, the collinear PrPsc, pairwisely in space draws together anticodons and forms a covalent and discrete “information RNA analog ” (iaRNA). This stage is practically a reverse process of the protein synthesis on a ribosome. The process is likely to take place on the ribosome’s A- and P-sites. Then, the synthesis of RNA on iaRNA takes place. For this purpose, a respective RNA polymerase, which can work with an iaRNA covalently-discrete matrix, is required. That’s the mechanism of “mutual usage” of the protein-synthesizing apparatus during the prion reproduction period. This impermanence creates the illusion that prions don’t possess a genetic apparatus. In this process, prion peptide chains are used as matrices on which poly-tRNA-continua in pairs arrange themselves on the ribosome’s A-P sections, forming discrete polyanticodons. The latter, joining in pairs , either become a direct matrix for the prion’s RNA-dependent mRNA synthesis, or (in the other case) polyanticodons may be specifically spliced and then alloyed in a covalently-undisrupted mRNA matrix of prions. Thus, prion’s mRNA polymerizes prions on a ribosome. That means that the ribosome operates in the reverse direction, being a “prion-polyanticodon-dependent mRNA polymerase” in the process. And, therefore, violating the dogma, information is transferred from a protein to RNA. Thus, the scheme of the DNA>RNA>Protein dogma completely changes. In this case, it isn’t the dogma any longer, but only a working model which needs further clarification and development. In accordance with this view on prion biogenesis, the prion stain-specificity is explained by peculiarities of reverse operation of ribosomes, temporarily recruited during the synthesis of each prion strain. These peculiarities reflect a taxonomic position of prion-producing biosystems. Now, back to the basic postulates of the genetic code model, still widely-accepted: the genetic code is a triplet, unoverlapped, degenerated and doesn’t have “commas”, i.e. codons are not separated from each other. Information flows from DNA through RNA to a protein. And finally, the code is universal. In light of the preceding arguments, what’s left out of the initial postulates? Nothing, essentially. Indeed, the code is likely to be a multi-letter fractal and heteromultiplet structure coding both individual proteins and functionally-linked protein associates. It has overlaps formed due to a shift in the ribosome’s reading frames. It has commas, since heterocodons can be isolated from one another by sequences with other functions, including punctuation functions. The code is not universal: in 14 cases, it is differed from the standard code of higher-level biosystems. The mitochondrial, yeast, micoplasm, trematodian and other lower organisms’ codes are included in these cases [5, 6].
And finally: a protein can be a matrix for RNA,
as we can see from the prion example. How should we understand
an actual protein code, taking into account all the
above-mentioned contradictions and in line with our theory? It
is possible to postulate a qualitative, simplified, initial
version of substance-wave control over the amino acids' line-up
order, dictated by the associates of aminoacylated tRNA, the
predecessors of proteins. Having admitted this assumption, it’s
easier to understand the operation of the protein code and
consider it as a hierarchically-structured program of the
substance-wave biosystem organization. In this sense, the code
is the first stage in a chromosome’s plan of building a
biosystem, since the genome language is multidimensional and
pluralistic and is capable of setting up more than just a
protein synthesis task. The basic statements of this proposed
preliminary model of matter-wave sign processes in protein
biosynthesis are as follows:
Multicomponent ribonucleoproteid protein-synthesizing apparatus is a system which generates highly organized sign radiation of acoustic-electromagnetic fields which strategically regulate its self-organization and the order of inclusion of amino acids in a polypeptide chain.
Aminoacylated tRNAs are assembled in sequences, which are the precursors of protein synthesis. This assembling is realized before the contact with the A-P ribosome site. The resulting continuum of tRNA anticodons pool is complementary to the complete mRNA, excluding dislocations determined by the availability of non-canonical nucleotide pairs.
The sequence of aminoacylated tRNA in associates-protein precursors is determined by the sign collective resonance of all the participants involved in the amino acid sequence synthesis. In this process, pre-mRNA and mRNA, which function as an integral continuum (macrocontext) of heteropolycodons variously scaled by length (including an intronic fraction pre-mRNA) are the key wave matrices. The main function of the wave matrices is an associative-context orientation of the aminoacylated tRNA sequence; this orientation works on a global scale, compared to F.Crick's “wobble-hypothesis”, superseding the rules of canonical pairing of nucleotides in the unidimensional space mRNA-tRNA. Laser-like radiations, emitted by the participants in this process and correcting the order of insertion of the amino acid components into a peptide, also function on the ribosome in addition to and/or together with the resonance regulations of a mutual dislocation of the codon-anticodon continuums. A ribosome enzymatically “de jure” fixes the peptide covalent bonds of amino acid sequences, selected “de facto” in a polyaminoacid-poly-tRNA-associate, the predecessor of the protein.
The resonance-wave “censorship” of the order of inclusion of amino acids in a peptide chain emends the potential semantic disorder in the creation of false protein “proposals” following from the homonymy of codon families, and ensures their correct “amino acid conceptualization” due to the context lift of the homonymy of multisided even doublets in codons The same mechanism is engaged in a higher-ranked ambiguity when the number of codons is (n+1).
Genetic code degeneration is necessary for pre-mRNA-mRNA-dependent, context-oriented exact matching of aminoacylated tRNAs, determined by the nature of wave associative resonance interactions in a protein-synthesizing apparatus.
The mechanism of generating the correct sequences of aminoacylated tRNAs on the wave matrixes of pre-mRNA-mRNA may be considered as a particular case of a partially complementary re-association of one-string DNA-DNA and RNA-DNA or, in general, as a self-assembly process known to characterize ribosomes, chromosomes, membranes and other molecular- and super-molecular cellular structures.
Ribosome can facilitate RNA synthesis on a protein matrix.
Thus, the role the mRNA plays is many-sided and dualistic. This molecule, like DNA, is a cornerstone in the evolutionary process and is marked by the mutually-dependent, synergistic unity of material and wave genetic information. An ambiguity of the material (substantial) coding is resolved by the precision of the wave information, which is likely to be realized through the mechanisms of collective resonance and laser-holographic (associative, contextual and background) effects in the cellular-tissue continuum. A jump to a more advanced level of wave regulation of the RNA-->Protein translation is accompanied by a partial or complete departure from the canonical laws of pairing of adenine with uracil (thymine) and of guanine with cytosine, which were attributable to the early (and simpler) evolutionary stages of DNA replication and RNA transcription. Such a refusal is informationally necessary, unavoidable and energetically preferable at a higher biosystem level. It’s worth stressing once again that the context associative-holographic mechanisms of operation of an organism’s protein-synthesizing system are tightly linked with the so-called “background principle” [44] and also with a multivector and multisided logic of a sophisticated system management (Gerhard Thomas’ kenogrammer) [26]. From this point of view, macrocontexts of pre-informational and contexts of informational RNA might be considered as a background which in this particular case is an “information noise source”. This permits to significantly amplify a signal under which the correct choice (wave identification) is made of one in two homonymous aminoacylated tRNAs, where only one of the two is to be incorporated in a protein correct “phrase”. This selection is only possible after a ribosome has managed to split a coherent component in the form of repeats of the same recognitions of one of the two similar doublets in codons. The following simplified example can explain the situation. Let’s suppose that it’s necessary to select one of two words (analogues of codons with doublets-homonyms). The words are “cow” and “bow”. It’s clear that the choice depends on the entire sentence, or on the context which helps to identify a signal, the correct word. If the sentence is “A good cow gives lots of milk”, then the replacement of “cow” with the word “bow” is equal to noise generation and to losing the signal. Pre-informational RNA and introns are likely to play a similar part; they are different levels of contexts which a live cell and its ribosome apparatus have to read and conceptualize to take a precise decision on tRNA anticodon selection in homonymy situation.
A family of various solitons (optical, acoustic, conformational, rotational-oscillating, etc.) excited in polynucleotides can become an apparatus for continual (non-local) “reading” of context RNA sequences on a whole. These solitons help gather semantic information on RNA contexts and then associatively regulate codon-anticodon sign interrelations. Genomes-biocomputers of cells carry out semantic estimates. Soliton reading, scanning the RNA surface, is a method of polynucleotide continual reading. For instance, the solitons of rotating torque vibrations of nucleotides on a sugar-phosphate axis we physically and mathematically considered for one-chain RNA-like DNA segments [30, 36]. These solitons respond to the nucleotide sequence alteration by the modulation of their dynamic behavior which acquires sign features and can probably be transmitted remotely, or over distances significantly exceeding the hydrogen bond length. Without a remote (wave, continual) migration of a signal containing information about the whole system, i.e. about pre-mRNA-mRNA-sequences, it isn’t possible to realize associative-context protein synthesis regulation. For this purpose, the wave capability of solitons (as well as of holographic memory) to deal both with separate parts and integral system as a whole, is required. This continuity or non-locality (which is the same) ensures that the ribosome apparatus recognizes and correctly chooses an actual codon from the two available doublet-homonymous ones, the codon, pseudo-noised with a background (context).
VII. Practical applications of genetic text linguistic ambiguities
What is the link between the above discussion and the problem of HIV and cancer research? Obviously, the link is direct. The HIV genome and oncogenes as well as other DNA structures, pseudogenes for instance, “are silent” (as factors of destruction), and this silence continues until a certain time. This key moment for initiation of a genome's pathological condition in cells, potentially inclined to abnormal development, is determined by transpositions of oncogenes and the HIV genome, or by transpositions of their polynucleotide surroundings in the chromosomal space and time structure. In both cases, the context environment of the pathogenic genome changes. The latter is no longer homonymous, unrecognizable or acceptable as a normal one by the cell. Other signals aimed at HIV reproduction are turned on (“are read and conceptualized”). A cell under the new context recognizes oncogenes as factors having other (pathological) command functions. The changed background (context) identifies and amplifies in the new polynucleotide situation potential signals and other meanings, which were hidden so far. The situation looks like that taking place in protein synthesis (choosing a correct codon out of the homonymic codons). Under this new context, cells are “confused in giving meanings” of DNA sequences and take-in wrong “decisions” as correct; this results in the complete shift of metabolism and its re-adjustment to a “cancer/viral way” - to reproduce the HIV genome. Here, a dualistic situation occurs: the new decisions are wrong in relation to the organism, but are right pertaining to the HIV reproduction. That’s how pathogens identify themselves and uncover their real “targets”, keeping and multiplying themselves as allogenic particles through the destruction of a biosystem as a whole. The problem of the DNA sequences migration in chromosomes may be discussed more globally (oncogenes, HIV genome or any other transposons whose purposes are still unclear for us). Moving along a genome as if over a context continuum, they obtain new meanings and other semantics which depend on their location in a 3D space of interphase chromosomes. The same logic is also true for “genetically-engineered” transgenesises of plants and animals. A growing number of artificial transgenetic organisms threatens with a global and rapid degeneration of all creatures living on Earth, because an uncontrolled automatic sign reconstruction of higher-ranked genetic codes, occurring after the introduction of foreign DNA molecules, isn’t taken into consideration. Practically uncontrolled intertaxonic transfer of foreign DNA-sequences, an avalanche-like semantic chaos in chromosomes and metabolic chaos in all biosystems (including human beings) will be the result of these genetically-engineered manipulations. It’s becoming hard to slur over the first alarming signals.
The rather abstract theoretical structures of genetic material transpositions we propose are confirmed not only by the example of transgenetic biosystems, but also by R.B.Hesin’s fundamental work [47]. Euchromatic genes, moving to an intercalar heterochromatin, produce a positioning effect, i.e. they are inactivated in some somatic cells and continue to function in others. Oncogenic cellular sequences are able to build-in in retroviral structures which didn’t originally have their own oncogenes. As a result, relatively non-hazardous viruses sometimes become tumorigenic. For instance, the RaLV rat virus might transform, having included master’s determinants in the genome, into the RaSV sarcoma virus. Cellular oncogenes, like viral ones, acquire a transforming activity if the lengthy repeated viral end sequences (LTR) are alloyed to oncogenes’ 5’-ends. In appropriate surroundings, proviruses including HIV viruses (as we think) are converted into latent (“silent”) genetic elements. They can persist in a master’s genome without producing any harm to it namely owing to the cellular DNA’s neighboring sequences repressing their activity. Taking into account this statement of Hesin’s, it’s possible to imagine a reverse situation, namely, the HIV genome activation in an environment of other DNA sequences when a cell in another DNA context already interprets HIV as a hostile semantic structure, but can do nothing to defend itself. However, as Mr. Hesin stresses, both peculiarities of the chromosomal DNA adjacent sections and operational principle which determine a provirus activity, are still a mystery. The mystery will remain unresolved, unless we apply new measurement criteria (semantically-vocal, wave or image measurements, i.e. the criteria we propose) to the genome. In this aspect, an interesting comparison of chromosome semantic and holographic information appears. A higher biosystem genome has several levels of information non-locality, “smearing” and redundancy, with a chromosome continuum holographic memory being one of them. Information locality and unambiguity of the genome’s mobile elements, the transposons, is contraposed to it; however, the multi-vector meanings of this information are developed dependent on a changing context of the transposon context surrounding; at the same time, transposons themselves are the triggers initiating the appearance, disappearance and repetition of the texts. A context “game” (combinatorial analysis) depends on current metabolic requirements of cells, tissues and an organism. The difference between a text and a context is conditional and depends on the domain of a part and an integer in a genome. The boundaries between the part and the integer are conditional and are likely to have a morpho-functional character which depends on an organism’s part differentiation at the cell, tissue, organ and biosystem levels. A finer ranking - by functional and metabolic areas of a cell which are controlled by certain chromosome sections (up to protein-genetic and exon-intronic splitting) - may also exist. Each of these quanta is an integral system in relation to itself, and just a part if the division rank is higher. Is it here that metabolic pathologies and herontologic manifestations are rooted when a biosystem stops identifying and differentiating many-sided patterns of a part and an integer? The HIV genome, like a transposon and a conditional part, might be invisible for a cell under some DNA contexts of master chromosomes . This is the way in which molecular-semantic mimicry of pathogenic chromosome structures is produced. Each coding-noncoding homonymous (or synonymous) and any other DNA sequence can be considered as a potentially multi-meaning pseudo-noised signal (signals) or as an image (images) which has to be identified and understood on the background of other dynamic gene images. The genetic apparatus amplifies each image signal and picks up the amplified signals out of the background (context, noise) not through the noise suppression procedure. On the contrary, a cell, a tissue and an organism use the background changing context as a means of extraction, amplification and to understand the meaning of each of these available image signals. It’s also logical to discuss in the same way the role of 3’- and 5’ - flanking sequences of protein genes highlighting one or another meaning. If we realized that the proposed mechanism of the dynamic game of genetic text meanings could play an important role in HIV and cancer development and in an organism’s entire metabolic status on the whole and if we accepted the idea that the comparison of a genome with natural texts and images wasn’t just a poetic metaphor, then real opportunities for the creation of a new biosystem management strategy, including management of viruses and oncogenes behavior, would emerge.
VIII. Is it possible to apply a probabilistic approach to identify individual (including pathogenic) meanings in a changing polysemantic genome continuum?
We have already mentioned some similarities between the Background Principle and Gerhard Thomas’ multi-vector logic (keno-grammar) and the prospects of these methodologies for the extraction and recognition of genetic or even metabolic vectors of multicellular organisms’ live functions. There’s one other direction in the natural languages theory, which, we hope, is applicable to genetic linguistic. This direction was developed by V.V.Nalimov and is linked with a probabilistic approach to understanding a language [22, 43]. V.V.Nalimov proposed that the semantics of any actual text (including a genetic one, we believe) could be described by its own distribution function (probability density), r (m ). Text revision and evolution are linked with a spontaneous manifestation of the filter r (y/m ), multiplicatively interacting with the initial function r (m ), in a certain situation y. We consider a “y-change” in a genetic text to be the natural transpositions of the DNA mobile elements, recombinations, the slicing and the alloying. The incorrect (for a biosystem) transpositions of its own (or foreign) DNA mobile elements, mutations and artificial transgenic manipulations are considered “unnatural changes”. An introduction of viral genomes, the HIV genome for instance, into a biosystem’s chromosome material, relates to a “specific class of unnatural changes”. The interaction of the r (y/m ) filter with the initial function r (m ) is ruled by known Buys’ formula:
r (m/y) = kr(m ) r(y/m ),
where
r (m/y) = distribution function determining the semantic of a new text after the “y-changes”
k = normalization constant.
According to V.V.Nalimov, Buys’ formula comes
forward as a syllogism: based on the two statements -
r (m) and r (m/ y), a text with a new semantic r (m/ y) comes to
life. Let’s assume that Buys-Nalimov’s logic is applicable to
genetic “texts”. Then the “idea” of these “texts” taken as a
whole is determined by 3 weight correlations which the r (m )
function specifies. “Meanings”, being a qualitative parameter in
nature, obtain a new quantitative characteristic. With the help
of the conditional distribution function r (m/y) V.V.Nalimov
presents a new interpretation, somewhat different from that used
in Buys’ statistics. In his theory, r (m/y) shows the
distribution density of a random value y under the given value m
. Therefore, not y, but m can be considered as an argument of
for the r (m/y) function which plays the role of a filter. We
think that the “y-changes” factor, initiating and exciting a new
semantic situation, is a key element in this model. Namely this
factor stimulates the unfolding of an increasing number of
alternative and new meanings, as well as of holographic
and other images in a variable semantic space of mobile DNAs in
a multicellular organism’s genome. The genome-carrying continuum
passes through the dynamic filter r (y/m) responding to it
by dramatic “y-changes”. It is significant that V.V.Nalimov had
been puzzled by the question of what permitted the reproduction
of the non-trivial r (y/m ) filters, but didn’t find an answer.
Nevertheless, at the same time he put forward an idea about the
role played by the environment and about a variety of situations
which could act as a source and a reason for adequate filter
formation. Here, V.V.Nalimov practically came up to the
above-discussed Background Principle. After the unification and
combination of Nalimov’s model and the Background Principle
statements it’s logical to consider that the y-factor is nothing
but a context (background) mechanism of switching on the r (y/m
) filters. These filters pick up the semantic loading and
meaning which are determined by an actual metabolic, including
genetic, situation: for instance - the necessity for a cell to
synthesize a huge amount of catalase at a given moment, a
process which is accompanied by a choice and the expression of
the catalase gene from a gene multi-meaning continuum. Herein
another, and perhaps the key mechanism of genome differential
activation to produce different proteins, is seen. Therefore,
the Background Principle and Buys-Nalimov’s logic became linked
by identical natural definitions. G.Thomas’ keno-grammar [26],
which is largely based on context orientations in choosing
priorities to manage complicated situations, is likely
convergent with the above-mentioned ideas.
Now back to the “genetic engineering”. Let’s also remind of the “chromosomal engineering”, when large blocks of a genome are used for production of useful hybrids. From the probabilistic approach to the mobile polysemantic chromosomal continuum, these “engineering” seem rather gloomy. Any manipulation here is an instant (as compared to the evolution pace) creation of new y-factors by people (and not by the evolution) and therefore, a mutation of the r (y/m ) purporting filters, unhampered by any time (evolutionary) frames. That’s the Earth’s genetic fund forthcoming chaos.
IX. The genetic apparatus paradox
The paradox of the genetic apparatus lies in the combination of two normally opposite properties - the stability of the information transferred from one generation to another, and the genome's volatility [47]. Genomic mobility is provided by polynucleotide transpositions, soliton-like non-linear dynamics (electric acoustic), and conformative and halogen restructuring. These non-random (programmed) movements of a chromosome continuum in live tissues are subtly and extensively distributed in a biosystem's space-time. The said dynamics is a means of the wave management of re-distribution of an organism’s various parts among each other. At the same time, it’s a method of metabolic event sequence organization. This strong sign chromosomal non-linear dynamics, which is easily found even in vitro, is realized through its isomorphous image in an organism’s space and time structure [32]. As a result, in a chromosomal continuum, as in a polysemantic and multiplex holographic formation, a permanent and variable semantic “game” of meanings goes on. Some kind of “endogenous semiotic show” of optical-acoustic regulatory (sign) images, which also have variable meanings, takes place. One of these chromosome images was experimentally found in many laboratories and is generally known as the phantom leaf effect (ref. to [32]). The phantom leaf effect theory is based on the principles of holography [32, 37]. It’s possible to say that the “game of meanings” is a function of sign dynamics of interphase chromosomes. This is a prerequisite for storing and processing vast volumes of information when a super-small volume of zygote mesomorphic chromosomes is able to operate a multi-vector and many-sided logic of development of extremely sophisticated biological systems. This is the origin of the idea that an essentially novel approach strategy to HIV and cancer treatment presumes the understanding and the possibility of managing a multi-vector genome logic. If we manage, applying genetic engineering methods, to purposefully and site-specifically introduce certain context DNA sequences to the 3’ and 5’ ends of oncogenes or HIV-genome, then it’s reasonable to expect the inactivation of their pathologic expression. On the other side, if we know the principles of ribosome operation in a context orientation mode, then we can successfully fight HIV in a ribosomal wave (laser, solitonic, polarization and radio wave) regulation zone. Ribosomes, synthesizing HIV proteins, must have thin wave vectors for management through context-background paths. Knowing them, it’s possible to suppress viral protein synthesis by external artificial modified fields similar to those normal cells use.
X. Genetic apparatus non-locality levels. Preliminary experiments.
Now, let’s turn to another genome operation phenomenon - that is, a supposed effect of quantum non-locality of chromosome sign conditions, which we have more or less experimentally confirmed [8, 37]. The idea of quantum non-locality was proposed by Einstein, Podolsky and Rosen [4] (EPR-effect). This effect is well in line with quantum physics experimental evidence. In short, the EPR-effect states that elementary particles, two photons for instance, which have initially been in a so-called “entangled” state, retain a mutual bond (this bond may be called “informational”) by quantum parameters (for example, by polarization), even if these elementary particles are removed from one another by any arbitrary distance. If the polarization of one of the particles has changed for any reason (for example, the photon passed through an optically-active layer and recorded the polarization modulations, then this photon disappears, but it manages to instantly transfer the recorded polarization information to another photon. To be more correct, it’s not a “transfer”, it’s a transition of one photon into another by means of a permissive teleportation mechanism. The first changed photon turns into the second one, independent of the distance between them. The second photon becomes a complete analogue of the first one. If this situation is in some way reflected in the genetic apparatus, then we rocket to new and higher orbits in understanding a metabolic processes and the Life phenomenon as a whole. In strictly physical terms, the EPR effect as a phenomenon of photon teleportation was correctly confirmed only in 1997 [2].
Other researchers soon obtained similar results, and not only based on photons. Multi-frequency physical fields are now teleported. Based on this data, it’s possible to suppose that photon fields, emitted by chromosomes as sign fields, can be teleported within or even outside the organism’s space. The same is true for wave photon fronts, which were read from the chromosome continuum similar to reading from a multiplex hologram. If photons are transformed into radio waves (the situation we found - ref. to [8, 33, 37]) through the EPR-mechanism, then this phenomenon is vital. In fact, the importance of quantum non-locality existence for a genome is hard to overestimate. We put forward and published this idea when we identified, with the help of the equipment we had developed, what was probably a more sophisticated variant of the EPR-effect. The said equipment includes a specially-designed laser which is capable of transforming its own photons into radio waves [46, 37, 8, 34]. The laser features a unique light beam dynamic polarization which could in some way simulate a dynamic polarization of chromosome laser radiations. It converts its photons (l =632.8 nm) into kHz-MHz-band radio waves upon the interaction of its beam with matter and the introduction of probing photons back in the laser resonator. Under these conditions, we suppose, pairs of entangled photons born in a gaseous phase of the laser optic resonator are transformed during their splitting and interaction with any body, including the laser mirrors, into radio waves. Photons were found to be able to localize in fractal clusters of the laser metal mirrors. If photons are probing an outer object, then the mirrors “store” its spectral characteristics. In such a way we have managed to record polarization & radio wave information of DNA preparations. This information carries morphogenetic signals. This fact enabled us to develop a fundamentally new type of dynamic polarization laser-radio wave spectroscopy and to investigate quantum non-local (teleportative) genetic processes.
We’d like to make some additional comments on the importance of quantum teleportation of genetic & metabolic information for biology on the whole. Quantum non-locality of genetic (chromosomal) information as a method of manifestation of its wave total distribution (continuity) in the space of multicellular biosystems seems to be just a particular case. In biosystems, there are at least six non-locality levels:
The first level is the constitutional (organism) level. Here, non-locality is expressed through the regeneration ability that organisms such as the planarium worms possess. After sectioning off any part, the worms’ body is able to reproduce an entire organism through regeneration. In other words, in this case there’s no link point between the genetic information common pool and a part of a biosystem. The same is also applicable to vegetative reproduction of plants.
The second level is the cellular level. It is possible to grow an entire organism from each cell (not only from a zygote). Despite the difficulties, it’s also possible for animal biosystems. Each cell is a potential continuum of an organism.
The third level is a cellular-nuclear level. Removal of nucleus from somatic and reproductive cells with a consequent introduction of another nucleus inside doesn’t impede a normal organism development. Such type of cloning has already been carried out at a higher biosystem level, on sheep for instance. Each nucleus of a cell is also a potential continuum of a biosystem. There’s no localization of genetic potencies at the level of individual cells.
The fourth level is a molecular level. The ribosome “reads” informational RNA either by individual codons, or on the whole, with the consideration of context, i.e. non-locally and continuously.
The fifth level is a chromosomal-holographic level. A genome possesses a holographic memory [37] which in nature is a typically-distributed (non-local) associative memory. At this and the next level non-locality obtains a new feature - a dualistic substance-wave character, since electromagnetic and/or acoustic fields, bringing out gene-wave information outside chromosomal material, “read” holograms as a substance. A physical field (or fields), marking the organism’s prospective space (calibration), comes on scene. The brain cortex holographic memory, establishing mental, semantic and image spaces calibrating potential actions of higher biosystems, is likely to belong to this category. In this way, social and genetic processes are alike.
The sixth level is a genome quantum non-locality. At the levels of up to 6th, genetic information non-locality is realized in an organism’s space. The 6th level is of a special nature, since it acquires a new quality. It’s manifested within the frames of one of the quantum non-locality forms, namely, in the permissive form we postulate in the current paper. In this case, non-locality is realized both by biosystem space and by its own, shrinkable to zero, time. Gene-wave programs, instantly spreading in such a way, simultaneously operate in an organism “here and there” and therefore, the semantic construction “now and then” loses its meaning. And this is a strategic factor and a vital evolutionary achievement of multicellular biosystems. Billions of organism’s cells have to instantly “know” a lot of information about each other. Without the “wave information instancy” phenomenon, a giant multicellular continuum of higher biosystems won’t be able to completely coordinate a metabolic process and its physiological and other functions. The intercellular diffusion of signal substances and nerve processes are too inert for this purpose. Even if we assume that sign electromagnetic fields are involved in an intercellular transfer process occurring with the speed of light (this assumption is quite reasonable), it’s not enough. A quantum non-locality mechanism, applicable to the genetic apparatus and which can act as an instantly-distributed quantum (wave) object isomorphous with substantial chromosomes, is required. Using non-locality, the genetic apparatus of higher biosystems creates an unparalleled phenomenon, where for certain intervals of time the “here and there” and “now and then” structures operate within the biosystems’ “closed” space-time as a continuity providing the organism with intrinsic super-coherence, information overredundance, a super-informativity and linkage and, as a result, proper integrity (survival). The ability of lower organisms’ (hydros, worms, amphibian, lizards, crustaceans) tissues and organs to regenerate (people have largely lost this ability ) is a manifestation of this phenomenon. But, considering the biosystem wave self-organization principles we are developing, it can be re-activated. The world’s first successful adaptation of donor tissues implanted to a blind man, which helped to return sight function to the patient, is a good example of regeneration. The principles behind this surgical operation and regeneration process is described in [33-35].
At the same time, theoretical and experimental research in this field is just emerging and needs further physical and mathematical understanding and development.
XI. Possible mechanism of recording information on laser mirrors
Now, let’s return to some features of the phenomenon of long-term recording of dynamic photon polarization-radio wave information on laser mirrors. We think this is linked with the phenomenon of photon fields localization (compression) in the system of correlated dispersers of laser mirrors. Given that the disperser material possesses a low radiation absorption ability, the external light field is capable to persist in the system for a long time without dissipation into other forms of energy. The reason for localization is connected with the interference of many times-diffracted waves. An external electromagnetic signal (in our case, it’s a laser beam modulated by polarization, for instance, by a DNA preparation) is localized (“recorded”) in the system of non-uniform laser mirrors. Later, the signal can be “read” without a significant loss of information in the form of isomorphously (in relation to photons) polarized radio waves. Theoretical research on a strain state of localized photons [12, 14-19, 24] seem to support these ideas. If this opinion is correct, then a chromosomal apparatus may also be considered as a fractal medium of localized photons accumulation, creating a coherent continuum with a quantum-nonlocally-distributed polarization radio wave genetic information. To some extent, this is in correspondence with our idea of genome quantum non-locality manifesting in one of its forms - ref. to [8, 34, 37]. It’s possible that the apoptosis phenomenon, which is likely to be involved in the regulation of multicellular creatures’ life time, is connected with an abnormal compression of photons by the nucleus of a cell, which are accumulated to a maximal value and then destroy the nucleus. The background principle of gene operation (including anti-oncogenes) may be another supplemental apoptosis regulation mechanism. For instance, an anti-oncogene coding the p53 protein could be controlled through the introduction of the DNA artificial flanking contexts from 3’-and 5’-ends of the p53 gene.
XII. Analysis of experimental evidences of gene wave forms existence
We are unaware (with some exceptions, of course) of modern publications on wave genetic theory and practice, as available in the major scientific journals. In the 1920-1940s, A.G.Gurwitch, A.A.Lyubitchev and V.N.Beklemishev, who developed the first theoretical models, were pioneers in this field; their ideas are described in detail in [32, 33]. In this paper, we are trying to produce more developed opinions of some possible synthesis mechanisms and functions of wave genetic structures, attributable to higher biosystems, as well as of the methods applicable for simulation of sign wave processes in chromosomes and model units simulating chromosome field functions and transferring wave genes. A publication and a patent, granted for the development of a device for the transfer of wave genes from a donor biosystem to an accepting one, are worth mentioning as an example of a rarely-appearing event. The said research was carried out by Yu.V.Dzang Kangeng [39, 40]. Kangeng’s device for a directed wave transmission of oncologic, including genetic, information to change hereditary characteristics of a biological accepting object is of a special interest. Unfortunately, there’s no theoretical interpretation of the device operation principles. Kangeng’s device has some common functional features with the equipment we developed and whose operation is based on similar principles. Kangeng’s device includes space elements (forms) which make it possible to split the radiation of a high-frequency SHF electromagnetic field generator into two orthogonally-polarized beams which repeatedly, as in our installation (in our case, it’s a laser beam transforming into radio waves), were passing through a donor biosystem and an accepting biosystem. Dzang Kangeng used a hexahedron, a cone, a sphere and a parabolic-reflector aerial as types of special forms. These forms provide a specific spinning (polarization) of the SHF (super-high frequency) field electromagnetic vectors. In our laser design, one of the mirrors used also had the form of a parabolic-reflector aerial directed to a resonator. During numerous repeated passes through an optically-active (an electromagnetic wave polarization rotating plane) hetero-liquid-crystalline donor biosystem, the organism’s tissues modulate the radiation (in our case, this is laser-radio wave radiation) by polarization, which is strengthened owing to repeated passes and is repeatedly and over a long time delivered to the accepting biosystem. In this process, the generator electromagnetic field “stores” the donor biosystem gene-sign polarization modulations in its “memory” then resonantly interacts with gene-sign polarization distribution of the accepting biosystem electromagnetic field. If the donor biosystem is at an early morphogenesis stage accompanied by an intense cell fission rate, it can’t be excluded that the supposed polarization resonances are also of a holographic nature. This many times-amplified signal, carrying the wave information that was “read” from the donor biosystem chromosome continuum, passes through the substance-wave structure of the accepting biosystem and makes it execute new gene-wave-polarization programs by means of the variation of their differential polarization structure. Changes in the accepting biosystem's gene-wave-polarization structure induced by the donor in the process of the field integration (“wave heterosis”) leads to a restructuring of its morphologic (genetic and phenotypic) characteristics. Shear wave correlations of polarization angles during the donor-accepting mixing of physical waves resulted in the acquisition of new morpho-genetic and biological properties from the accepting organism, are one of the most important quantum-electrodynamic events of the “wave hybridization” process. This fact allowed Dzang Kangeng with the help of the wave method to transfer genetic information from ducks to hens, for instance. Hybrid chickens of hens had typical features of a duck - a flat beak, an elongated neck, larger internal organs ( heart, liver, stomach and bowels). The weight of a one-year-old hen-duck hybrid was 70% higher than the weight of hens grown from irradiated eggs. The second generation of the hen-duck hybrids retained all changes, which were obtained in the first generation, even without further re-radiation. A wave transfer of peanuts’ features to sunflower seeds resulted in the change of form, taste and odor of a hybrid plant, which became similar to those of peanuts. Productivity grew by 1.8-fold; new features are transferred from one generation to another even without further re-radiation.
Let’s highlight some common features of the experiments Dzang Kangeng and we independently carried out: first and foremost, they demonstrate the possibility of genetic information existence in a wave form. This similarity is in the polarization modulation of the radiation orthogonal beams with intensity re-distribution in primary orthogonal beams with a frequency secured in the radio wave spectrum we register, by a donor organism. The spinning polarization planes here act as gene-semiotic structures whose biological meanings are identified and coded by angular and intensity shifts by a frequency spectrum. Similarly-polarized waves are known to be able to interfere, while orthogonally-polarized waves do not interfere at all. Waves with a partially-coinciding polarization produce, dependent on their polarization coincidence degree, a more or less sharp contrast interference picture. In other words, an angle cosine of each vector in relation to their registration plane or to the wave interference plane is a crucial factor.
Biology, including genetics and embryology, has already come to a turning point in its development, which is similar to the period when physics first admitted the idea that the properties of waves and particles didn’t contradict each other and were even complementary in quantum objects. A huge number of facts and scientific research outcomes available in modern molecular biology, genetics and embryology, can’t be understood without such a definition as physical fields, for instance, or without the application of quantum electrodynamics principles. The idea of lingual attributes of higher biosystems’ genome is a kind of humanitarian counterweight to an apparently excessive physical interpretation of basic Life function phenomena. The pace at which this idea is gaining acceptance by society is rather slow - in fact furious resistance has been encountered from certain circles. The current situation is easy to explain: the subject of Life is too complicated. Nevertheless, the time has come. If we are too late understanding the wave gene-sign functions of biosystems, it is possible that such diseases as cancer and HIV will destroy our society, or at least cause irreversible damage. We’ll also lose the opportunity for a mighty jump in biotechnology and biocomputing. Last but not least, we’ll also lose an opportunity to purposefully, rationally and positively influence sociogenetic and demographic processes. Following the above-described logic, we are coming to the conclusion that human speech structures, which provide the major information influx for mankind, possess fractally-scaled supergenetic properties. Evolution of society is similar to an organism’s morphogenesis. Books, libraries, movies, computer memory and people’s live speech in the end are the functional analogues of a cell chromosomal apparatus. The aim of these chromosomes is to control the creation of society space (houses, roads, oil- and gas pipelines, telephony, the Internet) and to arrange functional and structural relationships among the people inside it. Chromosomal sign properties, which have a lot in common with organisms, have a substance-wave nature. For instance, a movie showing an ideal model of a social structure and people’s relations within its frames is a substantial (material) formation (video tapes). However, it uses a mental-wave method to input information (light, sound, speech, idea, image). That’s the method chromosomes apply. The latter produce marking and calibration fields to arrange the organism’s space and also control information & metabolic relations, using, in particular, quasi-speech methods (let’s remember context orientations in the protein synthesis and function of oncogenes and HIV). Therefore, people ought to carefully study the operational principles of their own genetic apparatus and the “tricks” HIVs play to “mislead” our chromosomes. This kind of study is especially crucial today when Russia, and not only Russia, could face a demographic and social collapse within the next 5 to 10 years.
We have declared the theoretical approach to describe the logic of sign speech-wave relationships between HIV genomes and a master cell as well as the oncogene behavior logic. However, it’s not enough. We must obtain a set of key tools which will enable us to follow up at least the simplest wave command biocomputing functions of our chromosomes (1) and the reprogramming of our chromosomes by nucleotide sequences of HIVs and oncogenes. We have already developed this set of tools - it’s a laser uniquely reflecting coherent polarization-laser-radio wave (PLRW) quantum-non-local sign processes in chromosomes. Physico-mathematical formalism characterizing the PLRW-quantum processes in such appliances is presented in our research ( 2). PLRW-spectroscopy is the basis of wave information recording on laser mirrors - the phenomenon we have discovered. We have also managed to record information from specially prepared mesomorphic DNA matrixes, to broadcast it in a waveform at a distance of 1 m and to introduce it in accepting biosystems. As an accepting biosystem, we took plant seeds. Using this phenomenon, we effected a “wave reparation” of a genome of radioactively-damaged old seeds of Athaliana gathered in the Chernobyl Nuclear Power Plant area in 1987, and initiated drastic changes in stem and tuber phenotype in the second generation of the Solanum tuberosum plant. These biological influences don’t have the nature of mutations, they only have a sense meaning and are just another evidence that genetic information can exist in the form of electromagnetic fields.
No less important is the fact that genetic information can be recorded, stored, read, transmitted and introduced in accepting biosystems. Here, two vital factors emerge. The first one is that the recording of vast information volumes (including the genetic one) is an unparalleled event which confirms that it’s possible to develop principally new carriers of the dynamic super-capacity analog memory (images, texts). This is rather important for future biocomputing. The second factor is that owing to the PLRW phenomenon we enter a huge area of genetico-metabolic wave sign processes. Numerous and unclear events of distant “recognition” of the antigene-antibody and tRNA anticodon-iRNA codon pairs, as well as complementary mutual recognitions of DNA single chains, self-construction of ribosomes, recognition sites of ferments, careful piloting and landing of transposons in the DNA and so on, are also well contained within the frames of these processes. None of these phenomena can be explained by only Brownian movement and adjacent van der Waals, ion, hydrogen and electrostatic interactions.
And finally, the most important thing for us in the context of the ideas we propose is a wave and sign behavior of viruses, HIV or influenza, for instance. Viruses can be considered as “orphaned” cells which retained a minimum of chromosomal information required for a wave search of landing site on a master cell and exact place to cut-in own DNA as a transposon in the master cell’s DNA with consequent possible precise re-transpositions. Wave “languages”, which viruses use during the information contact with a cell’s surface and its genome, are the most vulnerable parts of a virus. Viruses use these “languages” to enter the semantic space of a cell and then to “mislead” the cell; after that, they undergo mimicry and are reprogrammed, reproduced and thus survive in the end. Cells are likely to be able to “mislead” viruses as well, creating a kind of “wave immunity”. That’s why a certain balance of powers in the fight exists; the said balance can shift in favor of a virus - for instance, in favor of influenza virus if the temperature starts fluctuating. Cooling of a blood circulation in nose mucosa capillaries changes the temperature of liquid crystals in chromosome blood cells. At the same time, protective wave programs recorded on high topologies of chromosome mesomorphic phases can only be slightly distorted. As a result, the “cold temperature information breach” appears and is used by influenza virus to reproduce. As a response to this action, a compensatory reaction evolves in the organism, i.e. the body temperature goes up to a sub-lethal level of 41° C. According to our thinking, this reaction is designed to “submelt” mesomorphic phases of the virus nucleic acid and, therefore, to produce noise or completely erase virus wave programs which it needs to attack the organism’s wave semantic space, thus to kill the growing number of its cells. Virus genome acoustic fields tightly linked with photon ones might act as wave bioprograms. Using the method of correlative laser spectroscopy, we demonstrated drastic changes in acoustic performance of the DNA liquid crystals in vitro at temperatures of 40-41° C; the results obtained partially confirmed our suppositions. And that’s only an example of wave sign processes in the relationship between the influenza virus and the human organism. Similar sense relationships exist between HIV and human cells, and the same issues arise - how to correctly find a landing site on a cell’s surface and precisely build-in the DNA (reverse transcriptasal copy of a viral RNA) as a mimicking transposon into the master cell DNA. Thereafter, the task is to get accurately re-transposed in a proper place on a chromosome and to detect and realize itself as a reproducing pathogen.
For now we can initially list the bottlenecks of HIV wave programs and name countermeasures to eliminate the problems:
Searching and recognition of HIV on a landing site (by altering the radiation nature of a virus and/or sites of landing on a cell, it’s necessary to distort the system of resonance-wave recognition mechanisms).
Searching and recognition of a viral DNA on the landing site by the master cell’s DNA (altering the radiation nature of a virus and/or sites of landing on the cell’s DNA, it’s necessary to distort the system of resonance-wave recognition mechanisms).
Searching and mutual recognition: protein's mRNAs of HIV«tRNAs (codon-anticodon recognitions) and proteins of HIV«RNA of HIV (self-assemble) for wave distortion of this process.
Any violation of even small wave sign resonances in this triad will result in the loss of infectious ability of HIV and other viruses, and Nature has created an example. As it was already mentioned, it’s an organism’s temperature mode. In ways similar to the one found by Nature it’ll become possible to design a simple “wave” vaccine against HIV and other viruses and bacteria. Our goal is to study the “alphabet” and “grammar” of wave “languages” of viruses’ genomes. And the foundation for this study has already been laid. A laser capable to “read” PLRW-wave genetico-metabolic information has been developed. However, the research in this field is rather difficult due to intrinsically natural inertia of the material understanding of genetic and metabolic information. Technical issues also exist. The laser we use generates only red photons, while the chromosomal apparatus of human beings and viruses uses a wide spectrum of coherent radiation ranged from 250 nm to 800 nm. Therefore, it’s necessary to design lasers which function in a full span of the spectrum visible area. This aim is technically feasible, but significant investments are needed to achieve it. In our opinion, all attempts to produce a material vaccine or other drugs to fight against the HIV or influenza virus will fail. Viruses continuously change their antigenic composition and thus bury all efforts of immunologists and other scientists engaged in the vaccine development. Efforts to chemically block certain stages of virus morphogenesis are inefficient and only poison human organisms. Wave vaccine is a reality. This vaccine would be non-invasive and environmentally-friendly, since it touches only a narrow area of wave sign relations between a virus and a cell.





